N2O reduction activity of isolate TF716
Strain TF716, which was isolated from the N2O-reducing consortium, was identified as Pseudomonas sp. (Fig. 1), which is the first N2O-reducing rhizobacterium to be isolated from the rhizosphere soil of tall fescue. Other N2O-reducing Pseudomonas spp. have been isolated from soil18,19, biofilters for treating landfill leachate20, and wastewater treatment plants21. Azospira spp. are N2O reducers that have also been isolated from wastewater treatment processes8,19,22. Other N2O-reducing bacteria that have been reported include Bacillus23 and Ralstonia24 isolated from soil, Paracoccus25 and Alicyliphilus22 isolated from wastewater treatment processes, and Aeromicrobium26 isolated from human fecal manure. Usyskin-Tonne et al.27 also isolated the N2O-reducing bacteria Agrobacterium/Rhizobium, Alcaligenes, and Pseudomonas from wheat root. Lee et al.12 reported N2O reduction by the bacterial consortium from the rhizosphere of maize and tall fescue.
Figure 2 presents the N2O reduction activity of isolate TF716 for different N2O loads (200–1500 ppm in the headspace). Isolate TF716 reduced N2O without a lag period under the experimental N2O loads (Fig. 2a), with the N2O reduction rate increasing with higher N2O loads until 750 ppm (15 µM in liquid) before slightly decreasing (Fig. 2b). Other studies have also reported that the N2O reduction rate increases with the concentration of N2O supplied to the final electron acceptor8,28. Based on the Lineweaver-Burk plot in Fig. 2c, the maximum N2O reduction rate (Vmax) produced by isolate TF716 was 18.9 mmol N2O·g dry cells− 1·h− 1. In past studies, the N2O reduction rate for Pseudomonas spp. has been reported to be 2.2–2.6 mmol N2O·g dry cells− 1·h− 1 8,19,21, while N2O-reducing Azospira spp. exhibited a N2O reduction rate of 1.60 − 23.85 mmol N2O·g dry cells− 1·h− 1 8,19,22, and that for Alicycliphilus denitrificans was 7.6 mmol N2O·g dry cells− 1·h− 1 22. Considering these results, the N2O-reduction ability of Pseudomonas sp. TF716 was superior to that of other N2O-reducing bacteria except Azospira sp. strain I13 (23.85 mmol N2O·g dry cells− 1·h− 1 )22.
Effect of the root exudate on N2O-reduction activity
Tall fescue and maize are widely used for the rhizoremediation of oil-contaminated soil because their roots are well-developed29–32. The interaction between the roots and rhizobacteria is a key parameter for rhizoremediation performance, with the root exudate potentially positively or negatively affecting rhizobacteria activity33–37. The effect of tall fescue or maize root exudate on N2O reduction by Pseudomonas sp. TF716 is presented in Fig. 3. Compared to the control (i.e., no root exudate), N2O levels were more rapidly reduced when the root exudate was applied. The N2O reduction rate for strain TF716 increased 1.5-fold in the presence of the root exudate, though there was no significant difference between the exudates for tall fescue (14.7 ± 0.3 mmol N2O·g dry cells− 1·h− 1) and maize (14.4 ± 0.8 mmol N2O·g dry cells− 1·h− 1). Greater N2O reduction with the addition of root exudate has also been reported in previous studies12,38. For example, when root exudate was added to N2O-reducing consortia, the N2O reduction rate was 1.3–2.7 times higher than without the exudate12. The root exudate of maize and tall fescue contains various organic compounds such as sugars, amino acids, organic acids, fatty acids, alcohols, and phosphate39–41. It is assumed that these organic compounds were utilized as nutrients or growth-promotion factors for strain TF716, thus leading to a higher N2O reduction capacity12,42.
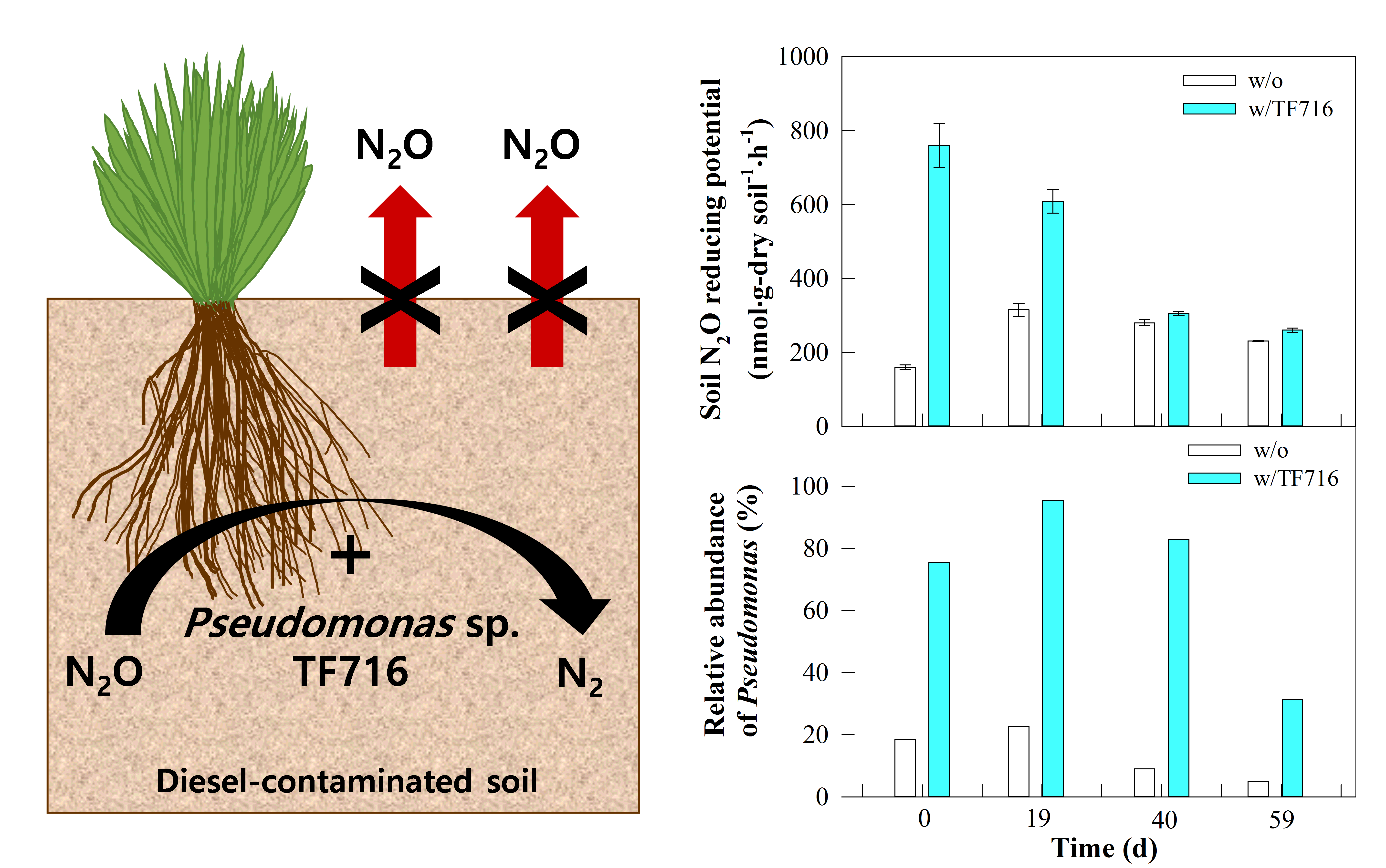
Inoculation effect of isolate TF716 on the N2O-reduction potential of the soil
Table 1 presents the inoculation effect of isolate TF716 on the N2O-reduction potential of diesel-contaminated soil planted with tall fescue for rhizoremediation. In the soil without the inoculation, the N2O-reduction potential was 159.4 ± 6.7 nmol·g-dry soil− 1·h− 1 on day 0, reached its maximum on day 19 (315.±17.4 nmol·g-dry soil− 1·h− 1), and decreased to 279.9 ± 8.6 and 230.6 ± 0.9 nmol·g-dry soil− 1·h− 1 on days 40 and 59, respectively. This could be because the relative abundance of those bacteria involved in N2O reduction increased after the planting of the tall fescue. However, when isolate TF719 was added, the N2O-reduction potential ranged from 609.1 ± 32.2 to 760.1 ± 58.9 nmol·g-dry soil− 1·h− 1 between days 0 and 19. On days 40 and 59, the potential was 304.8 ± 5.1 and 260.3 ± 5.8 nmol·g-dry soil− 1·h− 1, respectively, half the initial value. This indicates that the population of the inoculated strain remained dominant in the soil for 19 days, but its relative abundance subsequently decreased.
Table 1
Comparison of the soil N2O-reduction potential during the rhizoremediation of diesel-contaminated soil
|
Sampling time (d)
|
Soil N2O-reduction potential (nmol·g-dry soil-1·h-1)
|
|
TF
|
TF + 716
|
M
|
M + 716
|
|
0
|
159.4 ± 6.7i
|
760.1 ± 58.9a
|
159.4 ± 6.7i
|
760.1 ± 58.9a
|
|
19
|
315.1 ± 17.4c
|
609.1 ± 32.2b
|
211.5 ± 11.2hg
|
268.3 ± 15.1fde
|
|
40
|
279.9 ± 8.6cde
|
304.8 ± 5.1cd
|
239.5 ± 11.3feg
|
213.9 ± 14.6hg
|
|
59
|
230.6 ± 0.9fg
|
260.3 ± 5.8fdeg
|
177.1 ± 1.7hi
|
156.5 ± 2.4i
|
| TF: soil planted with tall fescue; TF + 716: soil planted with tall fescue inoculated with strain TF716; M: soil planted with maize; M + 716: soil planted with maize inoculated with strain TF716. |
When isolate TF716 was added, the N2O-reduction potential of the soil was 2.88 times higher than with no inoculation during the initial period (0 − 19 d) and 1.08 − 1.13 times thereafter. These results suggest that N2O emissions during the rhizoremediation of diesel-contaminated soil using tall fescue can be reduced by adding Pseudomonas sp. TF716. Usyskin-Tonne et al.27 reported that N2O emissions from wheat rhizosphere soil can be reduced by the addition of Agrobacterium/Rhizobium spp, while P. stutzeri PCN-1 can reduce N2O emissions during wastewater treatment43. In support of this previous research, the present study also demonstrates that the addition of N2O-reducing bacteria is a promising strategy to reduce N2O emissions during the rhizoremediation of diesel-contaminated soil.
In the soil planted with maize without isolate TF716, the N2O-reduction potential increased over the first 40 days from 159.4 ± 6.7 to 239.5 ± 11.3 nmol·g-dry soil− 1·h− 1, and then it decreased to 177.1 ± 1.7 nmol·g-dry soil− 1·h− 1 by day 59 (Table 1). This change in the N2O-reduction potential with maize was similar to that with tall fescue. When the soil with maize was inoculated with isolate TF716, the soil N2O-reduction potential decreased over time from 760.1 ± 58.9 on day 0 to 156.5 ± 2.4 nmol·g-dry soil− 1·h− 1 on day 59. Thus, with the addition of isolate TF716, the N2O-reduction potential increased 4.8- and 1.3-fold on days 0 and 19, respectively, but was slightly lower than without inoculation after 40 days. This indicates that the inoculated strain did not maintain its dominance in the soil planted with maize. Ferrarezi et al.44 investigated the inoculation effect of rhizobacteria on maize growth. Depending on the combination of the inoculating rhizobacteria, maize growth was either promoted or inhibited. Thus, when introducing a rhizobacterium, the interaction between the bacterium and the plant roots must be clearly understood to ensure that it is a positive combination.
Inoculation effect of isolate TF716 on rhizoremediation performance
Although isolate TF716 has no ability to degrade diesel itself (data not shown), the effect of its addition to the soil on diesel degradation was evaluated because other soil microorganisms may be affected by its presence (Fig. 4). In the soil planted with tall fescue, the addition of isolate TF716 increased diesel degradation compared to non-inoculated soil, though this difference was not significant. In the soil planted with maize, the diesel degradation in the non-inoculated soil (85 ± 0.8%) was slightly higher than in the inoculated soil (80 ± 0.4%) on day 19, but there was no difference on day 59. Overall, TF716 inoculation did not affect diesel degradation during rhizoremediation, suggesting that the isolate does not inhibit the activity of the microorganisms that are involved in diesel degradation.
In general, plant growth in contaminated soil is inhibited by pollutants, but it can be improved with the introduction of rhizobacteria. For example, the root growth of castor beans was promoted with the addition of a bacterial consortium into Pb and Zn-contaminated soil34. The addition of Burkholderia phytofirmans PsJ also reduced the inhibition of the root and stem growth of ryegrass in diesel-contaminated soil45, while the inhibition of mangrove in diesel-contaminated soil was reduced by introducing Bacillus, Sphingomonas, and Rhodococcus46. In addition, Gordonia sp. S2RP-17 enhanced the growth of maize in diesel-contaminated soil47, and the shoot and root growth of tall fescue in PCB-contaminated soil increased when the soil was inoculated with Bulkholeria spp.48.
To investigate the effect of isolate TF716 on plant growth, its indole-3-acetic acid (IAA)-production ability and 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase activity was first assessed, and it was found that it produced IAA but not ACC deaminase (data not shown). The inoculation effect of the isolate TF716 on the root growth of tall fescue and maize was then evaluated (Fig. 5). Compared with the control groups without inoculation, the addition of strain TF716 slightly enhanced the root growth of tall fescue, although this was not significant. In the soil planted with maize, strain TF716 had little effect on the N2O-reduction potential and diesel degradation (Table 1 and Fig. 4), but it unexpectedly enhanced the root growth of maize. Further research is required to elucidate the reason for this.
Dynamics of the bacterial community associated with N2O reduction during the rhizoremediation of diesel-contaminated soil
Figure 6 shows the dynamics of 12 genera involved in N2O reduction during the rhizoremediation of diesel-contaminated soil. Bacterial species belonging to Bradyrhizobium, Aminobacter, Mesorhizobium, Shinella, Paracoccus, Azospirillum, Alcaligenes, Castellaniella, and Pseudomonas have been reported to reduce N2O 49–61. Some also contain nosZ genes, which are responsible for the reduction of N2O to N252,54,57,59,62−65. Bordetella and Acidovorax have been reported to be denitrifying bacteria with nirS and nar/nir genes, respectively66–68.
Based on the analysis of the bacterial community dynamics, the dominant genera in the diesel-contaminated soils were Pseudomonas, Bradyrhizobium, and Mesorhizobium. Pseudomonas exhibited the highest relative abundance in all soil samples except for that for the soil planted with maize taken on day 59. In the soil planted with tall fescue, the relative abundance of Pseudomonas was 18.5–22.6% for the initial period (0 − 19 d), decreasing to 9.0% and 5.0% on days 40 and 59, respectively. In the TF716-inoculated soil planted with tall fescue, the relative abundance of Pseudomonas was initially 75.5–95.5% before decreasing to 31.2% on day 59. In the soil planted with maize, the relative abundance of Pseudomonas tended to decrease with time with or without the presence of TF716. In all soil samples, the relative abundance of Bradyrhizobium and Mesorhizobium increased with time, reaching 1.5–2.3% and 1.3–5.3% on day 59, respectively. In contrast, the relative abundance of Parcoccus, Alcaligenes, and Catenllaniella decreased over time.
Correlation analysis between key parameters
Figure 7 shows the correlation between TF716 inoculation, the N2O-reduction potential of the soil, the TPH-removal efficiency, plant type, and bacterial genera. The N2O-reduction potential was positively correlated with the addition of TF716 and the relative abundance of Pseudomonas (which includes strain TF716). The N2O-reduction potential was positively correlated with the addition of isolate TF716 and the relative abundance of Pseudomonas. Pseudomonas has previously been utilized as a biosource for the rhizoremediation of TPH-contaminated soil69–72. In particular, the introduction of Pseudomonas, which has plant growth-promoting traits such as ACC deaminase, IAA, and siderophore production, enhances plant growth, consequently improving remediation performance69,70. The positive inoculation effect of Pseudomonas in mitigating N2O emissions in agricultural soils has also been reported61,73. The present study also demonstrates that N2O emissions can be reduced during the rhizoremediation of TPH-contaminated soil using Pseudomonas TF716.
The TPH-removal efficiency had a close relationship with the relative abundance of Bradyrhizobium and Mesorhizobium, which were significantly correlated with maize. They also had a positive correlation with each other. Their capacity for N2O reduction has been demonstrated in past research49–51, 53,54. Bradyrhizobium can degrade various hydrocarbons such as chloroalkanes, chloroalkenes, and benzonate74, and Mesorhizobium is a known TPH-degrader75. Bradyrhizobium and Mesorhizobium were also the dominant bacterial groups in n-decane-degrading consortia76. As shown in Fig. 6, the relative abundance of Bradyrhizobium and Mesorhizobium increased during the rhizoremediation process. Based on these results, it is clear that they play a major role in TPH degradation and N2O reduction in diesel-contaminated soils planted with maize and tall fescue. Although Azospirillum is known to promote maize growth77, it had a positive correlation with tall fescue but not maize in the present study.
There were also some significant correlations among 12 bacterial genera in the soil samples. Pseudomonas sp. TF716 had a negative correlation with most of the bacterial genera. Aminobacter had a positive relationship with Shinella, Azosporillum, and Bordetella, while Shinella was positively related to Paracoccus, Azosporillum, Bordetella, and Castelaniella. Paracoccus exhibited a high positive correlation with Achromobacter, Alcaligenes, Bordetella, and Castelaniella. Achromobacter was associated with Bordetella and Castelaniella, while Alcaligens had a close relationship with Bordetella and Castelaniella, and the relative abundance of Bordetella was positively related to Castelaniella. Aminobacter has been reported to degrade 2,6-dichlorobenzamide78,79. In addition, Shinella, Paracoccus, Azospirillum, Achromobacter, Alcaligenes, Bordetalla, Castenella, and Acidovorax are known to decompose TPHs75,80−84. Although no statistical correlation was found, these bacterial groups are assumed to be involved in TPH degradation and nitrogenous compound metabolism, including N2O production and mitigation.
{kind=link}