Study cohort
In total, 38 individuals receiving SARS-CoV-2 vaccinations were recruited at the Freiburg University Medical Center, Germany. Of those, blood was collected from 31 individuals vaccinated three times with the mRNA vaccines bnt162b/Comirnaty or mRNA-1273/Spikevax, with 5 individuals receiving a fourth vaccination, without a SARS-CoV-2 infection history and 13 individuals with SARS-CoV-2 breakthrough infections after a third mRNA vaccination. Breakthrough infections were confirmed by positive PCR-testing from oropharyngeal swab. All 13 individuals with breakthrough infections included in this study had mild symptoms without respiratory insufficiency (according to WHO guidelines1). Characteristics of the participants are summarized in Supplementary Table 1 including the results of the HLA-genotyping performed by next-generation sequencing.
Ethics
Written informed consent was obtained from all study participants. The study was conducted in accordance to federal guidelines, local ethics committee regulations (Albert-Ludwigs-Universität, Freiburg, Germany; vote: 322/20, 21-1135 and 315/20) and the Declaration of Helsinki (1975).
PBMC isolation
PBMCs were isolated from venous blood samples collected in EDTA blood collection tubes by density centrifugation with lymphocyte separation medium (Pancoll separation medium, PAN Biotech GmbH). PBMCs were stored at -80°C until further processing. The cells were thawed in pre-warmed RPMI cell culture medium supplemented with 10% fetal calf serum, 1% penicillin/streptomycin, 1.5% 1M HEPES (all purchased from Thermo Scientific) and 50 U/mL Benzonase (Sigma).
Sequence alignment
Sequence homology was analysed in Geneious® 11.0.5 (https://www.geneious.com/) using Clustal Omega 1.2.2 alignment with default settings2. Reference genome of human ancestral SARS-CoV-2 (MN908947.3) was obtained from NCBI database. Genome sequences of SARS-CoV-2 variants of concern (VOCs) B.1, B.1.1.7, B.1.351, P.1, B.1.617.2, B.1.1.529 BA.1 and B.1.1.529 BA.2 were identified via CoVariants (https://covariants.org/). Spike epitopes in ancestral strain and all VOCs were aligned according to their homology on an amino acid level.
Peptides and tetramers for T cell analysis
Peptides were manufactured with an unmodified N-terminus and an amidated C-terminus with standard Fmoc chemistry (Genaxxon Bioscience). All peptides showed a purity of >70%. To generate tetramers, SARS-CoV-2 spike peptides (A*01/S865: LTDEMIAQY, A*02/S269: YLQPRTFLL) were loaded on biotinylated HLA class I (HLA-I) easYmer® (immunAware) according to manufacturer’s instructions. Subsequently, peptide-loaded-HLA class I monomers were tetramerized with phycoerythrin (PE)-conjugated streptavidin according to the manufacturer’s instructions.
In vitro expansion of spike-specific CD8+ T cells and assessment of effector function
1.5 x 106 PBMCs were stimulated with the spike protein-derived peptides A*01/S865 or A*02/S269 and anti-CD28 monoclonal antibody (0.5µg/mL) for 14 days in RPMI cell culture medium supplemented with rIL-2 (20 IU/ml, StemCell Technologies). At day 4, 7 and 11, 50% of the culture medium was exchanged with freshly prepared medium containing 20 IU/mL rIL-2. After 14 days, PBMCs were stimulated with peptides again, and stained for CD107a for 1 h at 37 °C to analyse degranulation. Subsequently, brefeldin A (GolgiPlug, 0.5 μl/mL) and monensin (GolgiStop, 0.5 μl/mL) (all BD Biosciences) were added and incubation continued for four more hours, followed by surface and intracellular staining with anti-IFNy, anti-TNF and anti-IL-2-specific antibodies. For calculation of the expansion capacity and to assess the cytotoxic capacity of the expanded cells, peptide-loaded HLA class I tetramer staining was performed together with intracellular staining of Granzyme B, Granzyme K, Perforin and Granulysin.
Magnetic bead-based enrichment of spike-specific CD8+ T cells
CD8+ T cells targeting spike epitopes were enriched as described previously3. In brief, 5 x 106 to 20 x 106 PBMCs were stained with PE-coupled peptide-loaded HLA class I tetramers for 30 min at room temperature followed by incubation with magnetic anti-PE microbeads. Subsequent positive selection of magnetically labelled cells was achieved by using MACS technology (Miltenyi Biotec) according to the manufacturer’s protocol. The enriched spike-specific CD8+ T cells were analysed using multicolor flow cytometry. Cell frequencies were calculated as previously described3. Of note, only samples with ≥ 5 virus-specific CD8+ T cells were included in subsequent analyses. Accordingly, the detection limit of virus-specific CD8+ T cells in this study was: 0.25 – 1 x 10-6, depending on the initial cell input.
Multiparametric flow cytometry for T cell analysis
The following antibodies were used for multiparametric flow cytometry: anti-CCR7-PE-CF594 (150503, 1:50), anti-CCR7-BV786 (G034H7, 1:50), anti-CD4-BV786 (L200, 1:200), anti-CD8-BUV395 (RPA-T8, 1:400), anti-CD8-BV510 (SK1, 1:100), anti-CD8-BV421 (RPA-T8, 1:200), anti-CD11a-BV510 (HI111, 1:25), anti-CD28-BV421 (CD28.2, 1:100), anti-CD38-APC-R700 (HIT2, 1:400), anti-CD39-BV650 (TU66, 33:1), anti-CD45RA-BUV496 (HI100, 1:800), anti-CD45RA-BUV737 (HI100, 1:800), anti-CD107a-APC (H4A3, 1:100), anti-CD127-BUV737 (HIL-7R-M21, 1:50), anti-EOMES-PerCP-eF710 (WD1928, 1:10), anti-Granzyme B-PE-CF594 (GB11, 1:800), anti-Granzyme B-BV510 (GB11, 1:50), anti-IFN-γ-FITC (25723.11, 1:8), anti-PD-1-BV605 (EH12.1, 1:50), anti-PD-1-PE-Cy7 (EH12.2H7, 1:200), anti-TNF-PE-Cy7 (Mab11, 1:400), anti-HLA-DR-BUV395 (G46-6, 1:200), anti-CD137-BV650 (4B4-1, 1:100), anti-CD95-APC (DX2, 1:100) (BD Biosciences), anti-BCL-2-BV421 (100, 1:200), anti-CCR7-BV785 (G043H7, 1:50), anti-CD57-BV605 (QA17A04, 1:200), anti-CXCR3-PerCP-Cy5.5 (G025H7, 1:33), anti-IL-2-PerCP-Cy5.5 (MQ1-17H12, 1:100), anti-Ki-67-BV711 (CD28.2, 1:200), anti-Perforin-PerCP-Cy5.5 (B-D48, 1:200), anti-Granzyme K-PE-Cy7 (GM26E7, 1:25), anti-Granulysin-APC (DH2, 1:100) (BioLegend), anti-TCF-1-AlexaFluor488 (C63D9, 1:100) (Cell Signaling), anti-CD14-APC-eFluor780 (61D3, 1:400), anti-CD19-APC-eFluor780 (HIB19, 1:400), anti-CD27-FITC (O323, 1:100), anti-KLRG1-BV711 (13F12F2, 1:50), anti-T-BET-PE-Cy7 (4B10, 1:200), anti-TOX-eFluor660 (TRX10, 1:100) (Thermo Fisher). To discriminate live cells from dead cells, fixable Viability Dye (APC-eFluor780, 1:400) from Thermo Fisher or 7-AAD from BD Biosciences (ViaProbe, 1:33) was used. To facilitate staining of intranuclear and cytoplasmic targets, FoxP3/Transcription Factor Staining Buffer Set (Thermo Fisher) and Fixation/Permeabilization Solution Kit (BD Biosciences) were used, respectively. Finally, cells were fixed in 2% paraformaldehyde (Sigma) and samples were analysed on FACSCanto II or LSRFortessa with FACSDiva software version 10.6.2 (BD), or CytoFLEX (Beckman Coulter) with CytExpert Software version 2.3.0.84. Further analyses of the data were performed using FlowJo version 10.6.2 (Treestar).
Dimensional reduction of multiparametric flow cytometry data
For dimensionality reduction, flow cytometry data was analysed with R version 4.1.1 and the Bioconductor CATALYST package (release 3.13)4. Initially, viable and tetramer-positive CD8+ T cells (or subsets of those) were identified using FlowJo 10 in two separate multiparametric flow cytometry panels (activation panel: HLA-DR, BCL-2, PD-1, CD137, Ki67, TCF-1, EOMES, T-BET, TOX, CD38, CD45RA, CCR7; differentiation panel: CD45RA, CCR7, CD27, CD28, CD127, CD11a, CD57, CXCR3, CD95, CD57, CD39, KLRG1, PD-1). To facilitate visualization of the dimensionality reduction by t-SNE and diffusion map analysis, cell counts were sampled down to at least 20 cells per sample, and marker expression intensities were transformed by arcsinh-transformation with a cofactor of 150.
Serum IgG determination
Detection of anti-SARS-CoV-2 spike-IgG was performed using Anti-SARS-CoV-2-QuantiVac-ELISA (IgG) from Euroimmun according to the manufacturer’s instructions (anti-SARS-CoV-2 S IgG; <35.2 BAU/mL: negative, ≥ 35.2 BAU/mL: positive).
Neutralization assay
Samples of vaccinated individuals and those with breakthrough infections were tested in a plaque reduction neutralization assay as previously described5. In brief, VeroE6 cells were seeded in 12-well plates at a density of 4 × 105 cells per well. Serum samples were diluted at ratios of 1:16, 1:32, 1:64, 1:128, 1:256, 1:512 and 1:1024 in a total volume of 50 μl PBS. For each sample, a serum-free negative control was included. Diluted sera and negative controls were subsequently mixed with 90 plaque-forming units (PFU) of authentic SARS-CoV-2 (either B.1, B.1.617.2 (delta) and B.1.1.529 BA.1 (omicron)) in 50 μl PBS (1,600 PFU/mL) resulting in final sera dilution ratios of 1:32, 1:64, 1:128, 1:256, 1:512, 1:1024 and 1:2048. After incubation at room temperature for 1h, 400 μl PBS was added to each sample and the mixture was subsequently used to infect VeroE6 cells 24 h after seeding. After 1.5 h of incubation at room temperature, inoculum was removed and the cells were overlaid with 0.6% Oxoid-agar in DMEM, 20 mM HEPES (pH 7.4), 0.1% NaHCO3, 1% BSA and 0.01% DEAE-Dextran. Cells were fixed 72 h after infection using 4% formaldehyde for 30 min and stained with 1% crystal violet upon removal of the agar overlay. PFU were counted manually. Plaques counted for serum-treated wells were compared to the average number of plaques in the untreated negative controls, which were set to 100%. Calculation of PRNT50 values was performed using a linear regression model in GraphPad Prism 9 (GraphPad Prism Software).
Statistics
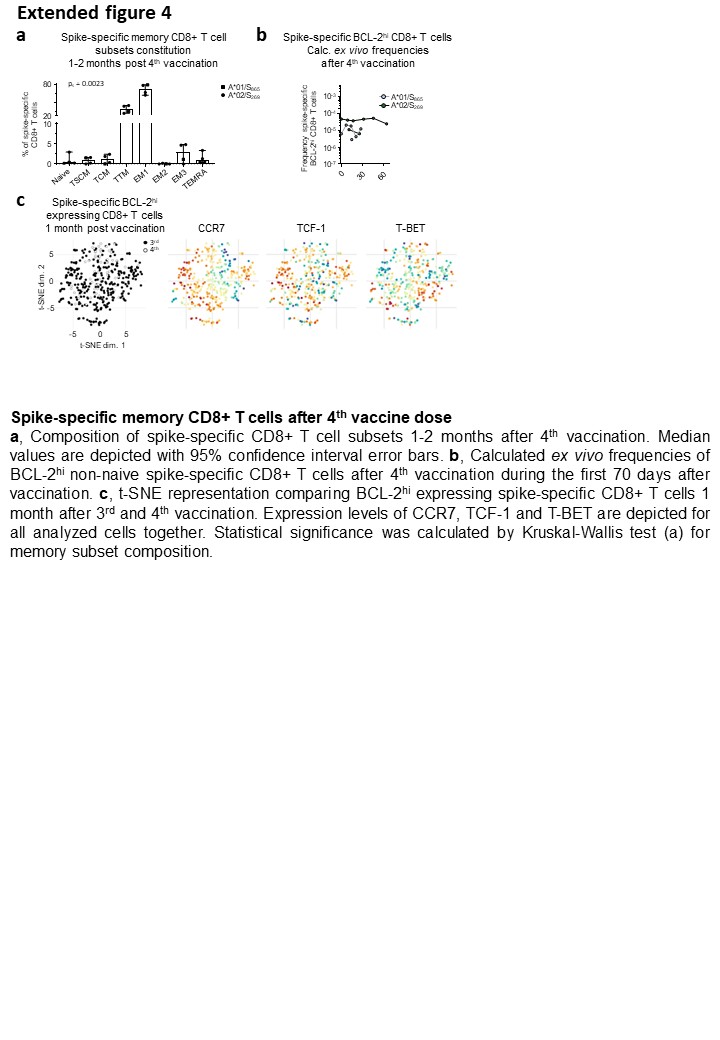
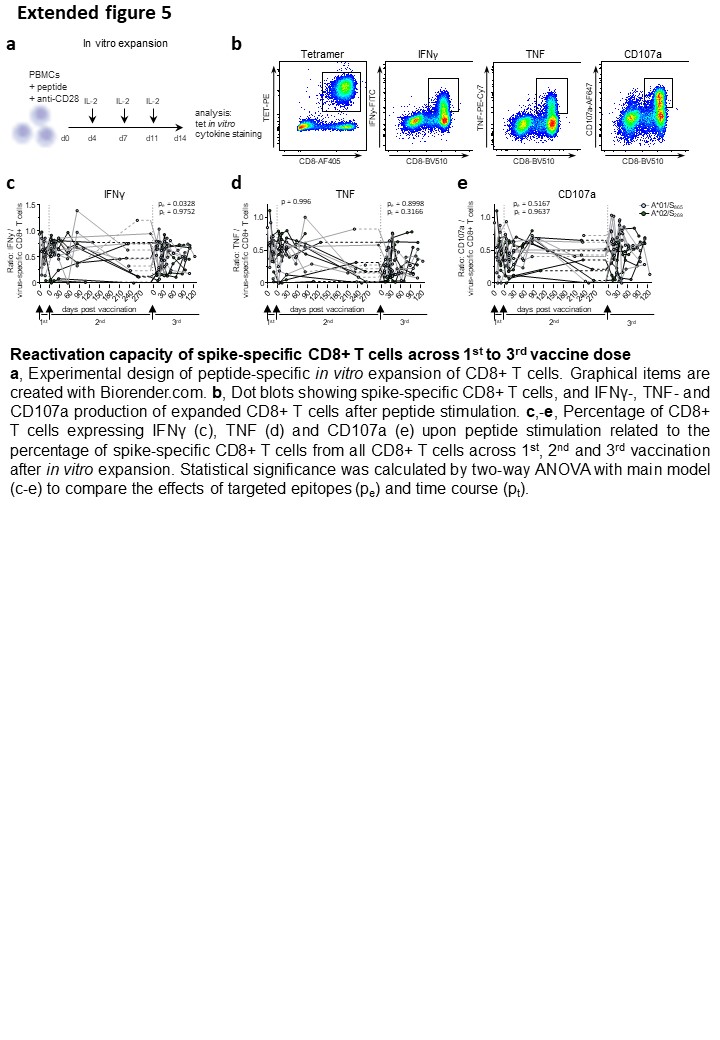
GraphPad Prism software version 9.3.1 was used for statistical analysis. Statistical significance was assessed by Kruskal-Wallis test, one-way ANOVA with mixed-effects model, two-way ANOVA with full model and main model. Statistical analysis was performed for A*01/S865 (n=7) and A*02/S269 (n=8) longitudinally analysed CD8+ T cell responses in Fig. 1A, 1B, 2C, 3A, 3B and Ext. Data. Fig 1C, 5C, 5D, 5E, for n=28 subjects longitudinally followed in Fig. 1D, for A*01/S865 (n=2) and A*02/S269 (n=3) T cell responses longitudinally followed in Fig. 1E, for n=26 subjects in Ext. Data. Fig 1G, for n=2 subjects in Ext. Data. Fig 2C, for n=7 at 3 months after 2nd vaccination, n=11 at 9 months after 2nd vaccination and n=11 at 3 months after 3rd vaccination in Fig. 2A and Ext. Data. Fig. 3B, for n=4 at 3 months after 2nd vaccination, n=8 at 9 months after 2nd vaccination and n=10 at 3 months after 3rd vaccination in Ext. Data. Fig. 3D, for A*01/S865 (n=7) and A*02/S269 (n=6) longitudinally analysed CD8+ T cell responses in Fig. 2D, for n=8 at 3 months after 2nd vaccination, n=12 at 9 months after 2nd vaccination and n=11 at 3 months after 3rd vaccination in Fig. 2B, for n=4 in Ext. Data. Fig. 4A, for A*01/S865 (n=2) and A*02/S269 (n=2) longitudinally analysed CD8+ T cell responses in Ext. Data. Fig. 4B, for n=10 at 3 months after 2nd vaccination, n=12 at 9 months after 2nd vaccination and n=11 at 3 months after 3rd vaccination in Fig. 3C, for n=10 at 3 months after 2nd vaccination, n=11 at 9 months after 2nd vaccination and n=11 at 3 months after 3rd vaccination in Fig. 3D, for n=6 at 3 months after 2nd vaccination, n=12 at 9 months after 2nd vaccination and n=10 at 3 months after 3rd vaccination in Fig. 3E, for n=6 at 3 months after 2nd vaccination, n=12 at 9 months after 2nd vaccination and n=11 at 3 months after 3rd vaccination in Fig. 3F, for omicron infection n=12, delta infection n=2 and 4th vaccination n=5 longitudinally analysed T-cell responses in Fig. 4A, for omicron infection n=11, delta infection n=2 and 4th vaccination n=4 analysed T cell responses in Fig. 4B and in peak response in Ext. Data. Fig. 6A, for omicron infection n=12, delta infection n=2 and 4th vaccination n=3 longitudinally analysed T cell responses in Fig. 4E, for omicron infection n=11, delta infection n=1 and 4th vaccination n=3 in Fig. 4F, for omicron infection n=6, delta infection n=2 and 4th vaccination n=4 analysed T cell responses after 1 month in Ext. Data. Fig. 6A and Ext. Data Fig. 6E, for omicron infection n=6, delta infection n=2 and 4th vaccination n=2 analysed T cell responses in Ext. Data. Fig. 6D.
Data availability statement
Source data files will be provided with this manuscript. All requests for additional raw and analyzed data and materials are promptly reviewed by the University of Freiburg Center for Technology Transfer to verify if the request is subject to any intellectual property or confidentiality obligations. Patient-related data not included in the paper were generated as part of clinical examination and may be subject to patient confidentiality. Any data and materials that can be shared will be released via a Material Transfer Agreement.
Code availability statement
R code to reproduce the analyses of multiparametric flow-cytometry data is available at https://github.com/sagar161286/SARSCoV2_specific_CD8_Tcells.
Online references
1 World Health Organization (WHO). Living guidance for clinical management of COVID-19. 23 November 2021
2 Sievers, F. et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol 7, 539, doi:10.1038/msb.2011.75 (2011).
3 Alanio, C., Lemaitre, F., Law, H. K., Hasan, M. & Albert, M. L. Enumeration of human antigen-specific naive CD8+ T cells reveals conserved precursor frequencies. Blood 115, 3718-3725, doi:10.1182/blood-2009-10-251124 (2010).
4 Crowell, H., Zanotelli, V., Chevrier, S. & Robinson, M. CATALYST: Cytometry dATa anALYSis Tools. R package version 1.16.2. (2021).
5 Oberhardt, V. et al. Rapid and stable mobilization of CD8+ T cells by SARS-CoV-2 mRNA vaccine. Nature 597, 268-273, doi:10.1038/s41586-021-03841-4 (2021).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}