Case study – Elraglusib first-in-human phase 1/2 study
A 55-year-old male patient with widely metastatic melanoma with involved organs including the lungs, bones, muscles, stomach, lymph nodes, pancreas and adrenal glands, as well as over 25 distinct brain lesions, was enrolled on the elraglusib 1801 first-in-human phase 1/2 study (EudraCT #:2018-003739-32; NCT #: 03678883). The patient’s melanoma was BRAF V600E mutated, and he had received multiple prior treatments including PD-1 inhibitors, CTLA-4 inhibitor, as well as BRAF/MEK inhibitor combination therapy. The most recent treatment regime prior to study entry was a combination of a PD-1 inhibitor with both BRAF and MEK inhibitors, the patient had progressive diseases on this regimen. The patient was without treatment, “washout-period”, prior to elraglusib study entry during which time there was particularly rapid progression of brain lesions (Fig. 1). Treatment began with single agent elraglusib at 5mg/kg IV given twice a week on days 1 and 4. After 6 weeks, MRI showed > 30% decrease in size in all brain lesions, and PET scan demonstrated an excellent response with near complete resolution of all tumours and only residual focus of uptake in the stomach, with no new lesions. After completing 12 weeks of elraglusib treatment, MRI showed 8 cystic brain lesions with no change in size; no new lesions and was considered a complete response by Response Assessment in neuro-oncology (RANO) criteria(30). PET scan showed no areas of residual uptake, as well as resolution of uptake in the stomach, and were consistent with a complete metabolic response to therapy. The patient remains on elraglusib single agent therapy and is now over 36 months in continuous complete response.
Evaluation of elraglusib for pre-clinical studies in mice
Previous work has shown that inactivation of GSK- 3 through small molecule inhibitors can enhance antigen-specific CD8 + CTL responses against tumour targets in vitro (9). Here we evaluate the use of small molecule drug, elraglusib, to enhance these CTL responses. Initially we examined responses of CTLs generated from OT-I transgenic mice that carry an MHC class I-restricted T cell receptor (TCR) specific for the SIINFEKL peptide of OVAlbumin (OVA257 − 264) as presented by H-2kb. OT-I T cells were activated by OVA257 − 264 alone or in the presence of elraglusib and/or anti-PD1 mAb for 5 days, following which the CTLs were subjected to a 4 hour incubation with EL4 lymphoma target cells before lactate dehydrogenase (LDH) release was measured as an indication of cytolytic killing. EL4 cells were pulsed with OVA-peptide prior to the incubation to ensure a specific response and non-pulsed EL4 cells were used as a control for spontaneous (background) killing. Three different doses of elraglusib were compared in combination with or without anti-PD1 mAb treatment. LDH release showed a significant increase in target killing in the presence of elraglusib with the concentration for optimal target death being determined as 10uM (Fig. 2A). At an effector to target ratio of 10: 1, elraglusib increased the percentage of target killing from 10–39%, at 25:1, killing increased from 32–64% and at 50:1 from 59 to 86%. CTLs treated with anti-PD1 mAb showed an increase in CTL activity compared to non-treated CTLs but had no further effect on CTLs treated with elraglusib at any of the doses used.
As predicted from previous work using GSK-3 SMIs, the upregulation of PD-1 expression due to OVA activation was dramatically reduced in the presence of elraglusib as demonstrated through PCR analysis (Fig. 2B) and flow cytometry (Fig. 2C). Although the combination of anti-PD-1 with elraglusib did not show any significant increase in cytolytic killing in vitro compared to the use of elraglusib alone, the combination did reduce PD-1 transcription at suboptimal levels (1 and 5uM) of elraglusib as seen by PCR and flow cytometry.
Elraglusib suppresses subcutaneous B16 melanoma growth as a single agent and enhances anti-PD1 therapy when given sequentially
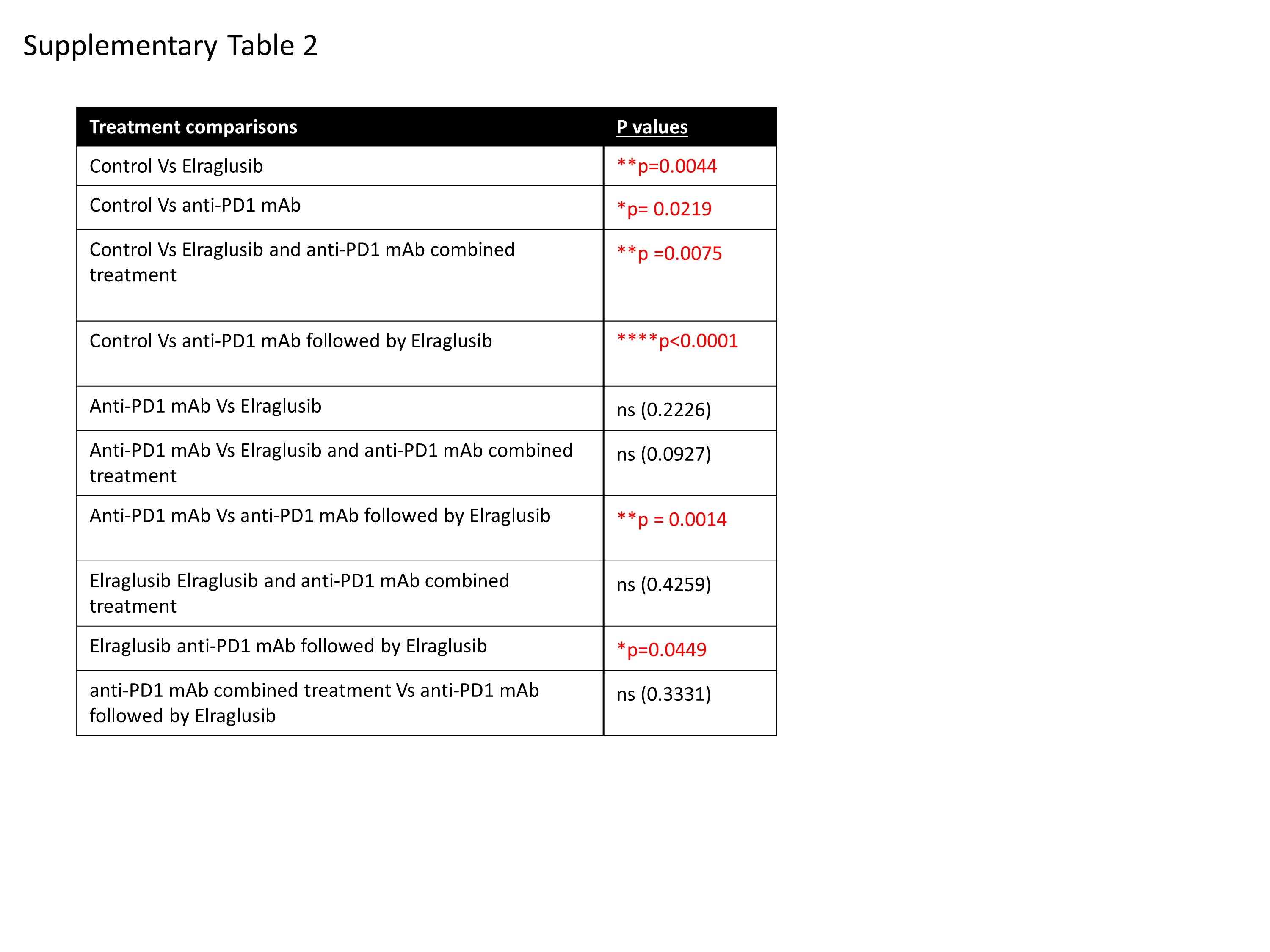
As elraglusib clearly enhanced cytolytic killing by CD8+ CTLS in vitro, we next assessed whether elraglusib could regulate tumour growth in an in vivo setting. To do this, B16 tumour cells were injected subcutaneously into C57/BL6J mice, followed by intra-peritoneal injections of either anti-PD1, elraglusib or a combination of the two as outlined (Fig. 3A). To mimic the findings found previously in the clinical case (Fig. 1), one group of mice received anti-PD-1 treatment and following a 48h break, went on to receive elraglusib treatment on days 13, 15, 17, 19, 21 and 23. Due to the aggressive nature of B16 melanoma tumour growth it was not possible to lengthen this “break” time between treatments. Tumour growth was monitored over time and survival was based on tumour burden, with mice sacrificed upon tumours reaching a maximum diameter of 15mm. Kaplan-Meier survival plots demonstrated increased survival in mice treated with elraglusib alone (50%) which was higher than that with PD-1 Ab treatment alone (30%) when compared to 0% of mice non-treated mice after 18 days post tumour cell injection. Moreover, this survival was increased to 80% when treated with a combination of both elraglusib and anti-PD1 Ab, and 100% when mice received sequential treatments (Fig. 3B). This increased survival was due to a decreased rate in tumour growth as can be seen from the daily tumour measurements (Fig. 3C). Flow cytometry demonstrated this suppressed tumour rate to be associated with a decrease in PD-1 expression when comparing tumour infiltrating lymphocytes (TILs) from the different treatment groups. Furthermore, this reduced PD-1 expression was primarily associated with elraglusib treatment and was further reduced due to combination or sequential treatments (Fig. 3D). Tumours and spleens were harvested 24 hours after the last treatment was given (day 14 for mice receiving elraglusib alone and or in combination with anti-PD-1, the samples for the sequential treatment group were taken on day 24, although there may be differences due to development of the tumour itself, it was decided to compare samples after completing treatment; to take samples from the sequential group at day 14 would not take into account the second part of treatment and would resemble those receiving anti-PD1 alone and to take all samples at day 24 was not viable due to low survival and any effects due to the treatment may no longer be apparent).
This effect on PD-1 expression was shown further through qPCR of isolated spleen T cells showing a marked decrease in pdcd1 transcription (Fig. 3E). Furthermore, this was accompanied by a decrease in other co-inhibitory checkpoint molecules LAG-3 and TIGIT. C-X-C Motif Chemokine Receptor 3 (CXCR3) on the other hand showed an increase in transcription due to the presence of elraglusib treatment.
Elraglusib as a single agent or in combination with anti-PD1 therapy significantly supresses the growth of intracranial B16 melanoma
Melanoma is well known to spread to various locations in the body, particularly to the brain and the B16 melanoma mouse model has been previously utilised to demonstrate this (31). Here we injected 2 x 105 fLuc tagged B16 cells intracranially into C57/BL6 mice and following the treatment regime shown (Fig. 4A), we tracked tumour growth over time using IVIS live imaging. Survival was based on when mice started to show initial symptoms of a brain tumour i.e., severe lethargy, weight loss, disorientation. Non-treated mice showed a maximum of 11 days survival following tumour injection which was increased to 60% of mice treated with elraglusib alone, 20% with PD-1 Ab alone and 50% when treated with a combination of both elraglusib and anti-PD1 Ab. However, this survival was increased further (80%) when mice were sequentially treated with PD-1 Ab followed by elraglusib (Fig. 4B). At day 8; (or day 14 for the sequential treatment group) 24 hours after the last treatment was given, mice were injected intraperitoneally with luciferin and scanned by IVIS Lumina imaging (Fig. 4C). Total flux (photons per second) was measured showing a clear reduction in luciferase signal in response to treatment with elraglusib. This is representative of the haematoxylin and eosin (H&E) performed at these time points (Fig. 4D). The control (DMSO vehicle treated group) shows the intracranial tumour to have a rapid growth rate with an invasive growth pattern as seen by the heavy staining. Both monotherapies show a more compact tumour area with the elraglusib showing a clear decrease in tumour size. Although the combination of elraglusib with anti-PD1 did not appear to significantly reduce the signal as seen by ivis, the H & E does show a clear decrease in tumour size which aligns with the increase in survival as shown in Fig. 4B. This is similar for the sequential treatment; in this case the tumour is more spread than in the combined treatment however the intensity of the staining is lower. It should also be noted that this sample was taken at day 14 which is 6 days later than the other samples, which clearly supports the increased survival seen in Fig. 4A. Quantitative PCR (Fig. 4E) on these samples revealed decreased transcription of checkpoint molecules, PD-1, LAG-3 and TIGIT as seen previously in the subcutaneous model (Fig. 3E), which was accompanied by an increase in CXCR3 transcription. This reduced expression of PD-1 was also seen in CD8 + TILs isolated from brain tumours in response to all 3 treatments incorporating elraglusib as demonstrated by flow cytometry (Fig. 5A). This was further confirmed through immunohistochemistry of sections taken from the samples in Fig. 4D. Staining showed CD8 + and PD1 + cells to be scattered throughout the tumour region (Fig. 5B) however, significantly decreased numbers of PD1 + cells were seen in all treatments (Fig. 5C), although there was no significant change in the number of CD8 + cells.
Elraglusib Significantly Improves Survival When Tumour Burden Is Spread Over Two Locations
In human melanoma, the majority of brain metastases coincide with extracranial metastases, usually present in the skin, to mimic this we utilised a 2-stage model as demonstrated by Taggart et al (39). In this model mice were initially given a subcutaneous tumour cell injection, followed by an intracranial tumour cell injection 3 days later. The kinetics of this and the therapy given were confirmed by the previous intracranial and subcutaneous studies and is summarised in the schedule shown (Fig. 6A).
Survival was based on when mice started to show initial symptoms of a brain tumour i.e., severe lethargy, weight loss, disorientation, or when the subcutaneous tumour reached 15mm in diameter. The previous experiments in Figs. 3 and 4 determined a mean survival time of 16 days and 10 days for animals receiving subcutaneous or intracranial tumours alone, respectively. Therefore, staggering the implantations allowed for quantification of intracranial tumour-dependent survival. Survival (Fig. 6B) is depicted as days post the initial subcutaneous tumour injection and therefore when comparing to the intracranial injections alone a difference of -3 days should be taken into consideration. Non-treated mice showed a maximum of 15 days survival following subcutaneous tumour injection which was increased to 25% with PD-1 Ab alone and 87.5% of mice treated with either elraglusib alone or as a sequential treatment. However, survival was only slightly increased to 12.5% when treated with a combination of both elraglusib and anti-PD1 Ab. This demonstrated the single agents in this case to be most effective, particularly elraglusib, and that giving simultaneous although slightly better than no treatment was detrimental compared to giving single or sequential treatment. Further to this the two different locations were assessed for the impact of the treatment on tumour growth. Live imaging (Fig. 6C) at day 10 (day 16 for those receiving sequential treatment) post subcutaneous tumour injection showed the intracranial signal to be significantly lower in both the elraglusib and the sequential treatment group supporting the increased survival of these groups.
Although all mice were subjected to two tumours, it can be seen from the growth curves (Fig. 6D) that survival of the majority of mice receiving DMSO (vehicle control) or PD-1 therapy either as a single agent or simultaneously with elraglusib treatment, was determined by the intracranial tumour, as the subcutaneous tumours in these animals did not reach the maximum 15mm in diameter. However, the majority of animals receiving elraglusib treatment alone (5 out of 8 mice) or as a sequential treatment (7 out of 8 mice) survived the intracranial tumour and succumbed to the subcutaneous tumour growth. 62.5% of animals (5 out of 8) receiving sequential treatment were surviving at day 19, which was 16 days following intracranial injection. This survival was markedly increased compared to mice receiving only the intracranial injection (Fig. 4B), which demonstrated 30% survival at day 16. A similar, albeit lower, trend was seen in those receiving elraglusib treatment alone (25% versus 10%).
Quantitative PCR (Fig. 6E) on these samples revealed decreased transcription of checkpoint molecules, PD-1, LAG-3 and TIGIT, accompanied by an increase in CXCR3 transcription. This reduced expression of PD-1 was also seen in both CD8 + TILs isolated from intracranial and subcutaneous tumours in response to all 4 treatments as demonstrated by flow cytometry (Fig. 6F)
{kind=link}
{kind=link}
{kind=link}