Idiopathic IBD, which includes CD and UC, creates several diagnostic impediments owing to common clinical, radiographic, endoscopic, and histologic characteristics. Thus it is important to recognize the subtypes of IBD for developing specific diagnostic and novel therapeutic intervention in IBD. The miRNAs are potential candidates for therapeutic application owing to well-defined mechanism of action during dysregulation in human IBD and simple oligonucleotide design [19]. This paper investigated the expression pattern of miRNAs in IBD in comparison to the other IBD subtypes, namely CD and UC, in order to identify miRNAs as potential diagnostic and prognostic biomarkers to better delineate the IBD subtypes.
In the present study, an analysis of the GSE53867 expression dataset revealed 16 DEMs (P value <0.05 and |log2FC| >1), including 10 upregulated and 6 downregulated DEMs, in IBD and its subtypes. Among the 16 DEMs, 3 miRNAs including miR–891a–5p, miR–27a–5p, miR–1204 were unique to the CD group, 2 miRNAs such as miR–342–3p, miR–16–5p were unique to the UC group, 4 miRNAs including miR–523–3p, miR–422a, miR–665, miR–519c–5p belonged exclusively to the IBD group. However, the common miRNAs under the UC and IBD were miR–1973, miR–142–3p and those under CD and IBD were miR–24–2–5p, miR–650, miR–24–1–5p, miR–491–3p; no common miRNAs belonged concomitantly to both UC and CD groups. This finding supported the presence, in the colon tissues, of miRNAs: miR–891a–5p, miR–27a–5p, miR–1204, indicative of CD; miRNAs miR–342–3p, miR–16–5p could be applied as a biomarker to identify UC; miRNAs including miR–523–3p, miR–422a, miR–665, miR–519c–5p could implicate the possibility of IBD diagnosis, while the presence of common miRNAs within CD and IBD, UC and IBD could be suggestive of overlapping epigenetic modifiers targetable by combined therapeutic applications. Occurrence of a similar few signature miRNAs included elevated miR–16 and miR–24, decreased miRNA miR–650, in mucosal tissue of UC; similar miRNAs miR–16 and miR–422a have been found significantly upregulated in the mucosal tissue of CD compared to normal healthy controls [20–22]. The panel of microRNAs that have been found differentially expressed between UC and CD in colonic tissue included miR–19b, miR–23b, miR–100a–3p, miR–100b–5p, miR–106a, miR–150, miR–191, miR–196b, miR–223, miR–320a, and miR–629 [23]. The colonic microRNA panels reported by different studies attempting to delineate UC, CD, between UC and CD exhibited little overlap, due to discordance in the activity status of IBD during sampling, location of tissue, inflammation level, previous medication, platforms for micro-RNAs estimation [24].
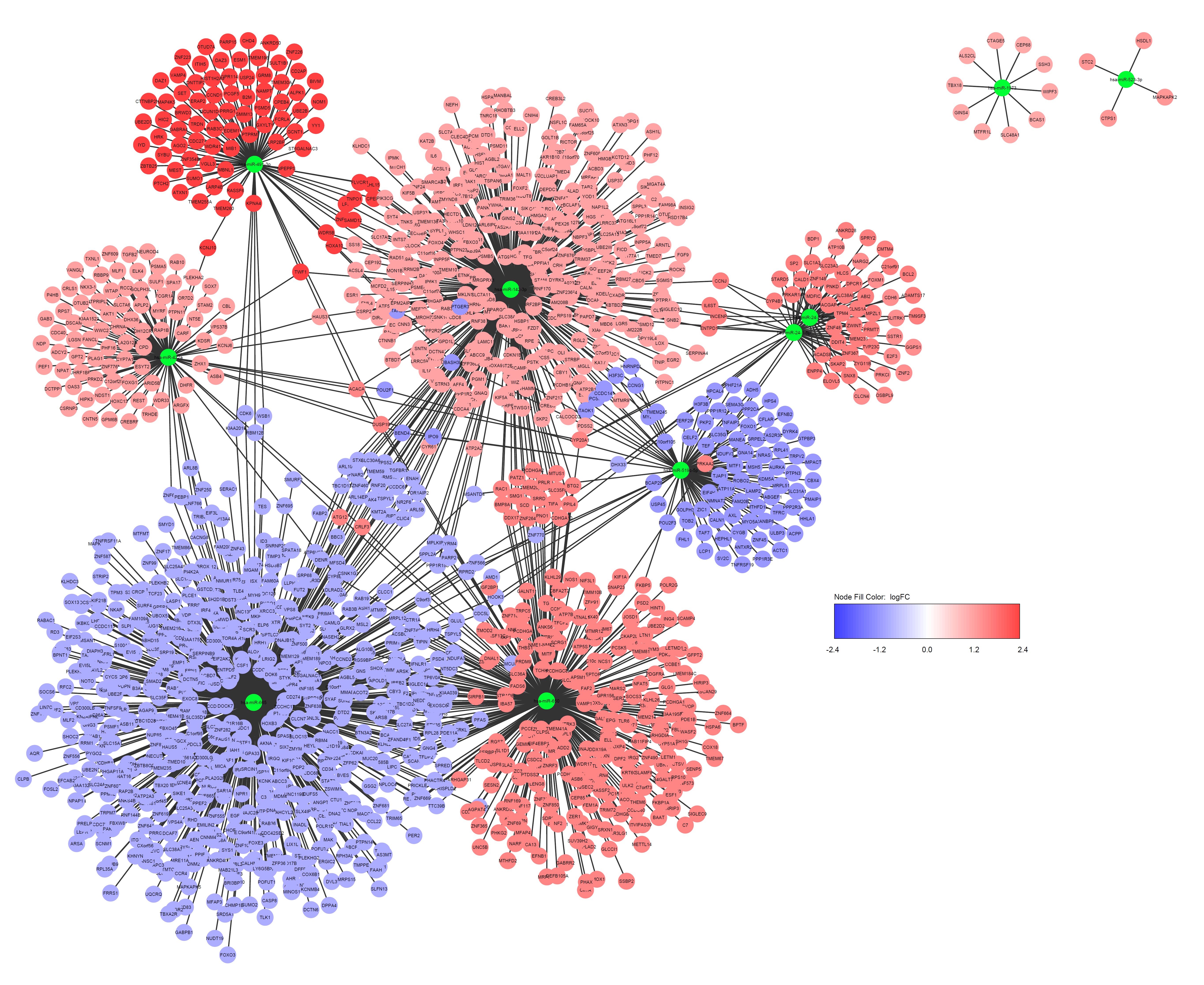
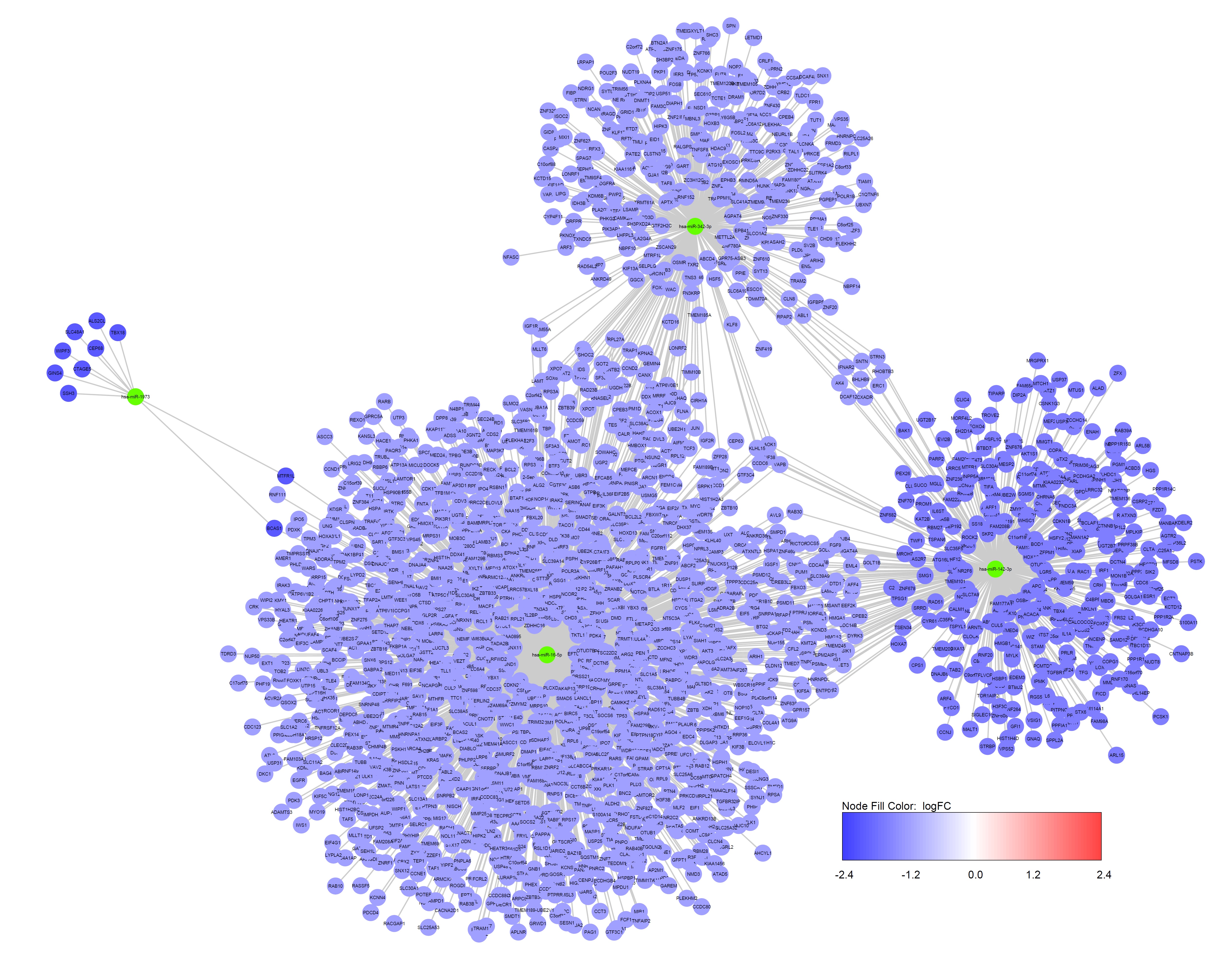
In the current study, genes which concurrently fitted 5 databases (miRWalk, miRDB, miRanda, RNA22, and TargetScan) were chosen as predicted target genes, and validated target genes from miRWalk were taken, amounting to 1673 upregulated and 3145 downregulated genes. The miRWalk database contains information about predicted interactions based on integration of several algorithms and experimentally validated interactions as well (http://mirwalk.umm.uni-heidelberg.de) [10].Selecting interactions predicted by multiple tools ensures enhanced recovery of identical miRNA-gene interactions. Cytoscape tool was used to visualize the interaction data on the predicted and validated miRNA and their putative target gene regulatory network structure [15]. There were 620 DEGs identified in the CD group targeted by 6 upregulated DEMs including miR–24 (n = 98), miR–650 (n = 341), miR–491 (n = 92), miR–891 (n = 14), miR–27 (n = 62), and miR–1204 (n = 12) (n = DEGs) (Suppliment Fig. 1S1); 2377 DEGs identified in the UC group targeted by 4 downregulated DEMs including miR–1973 (n = 10), miR–142 (n = 409), miR–342 (n = 358), and miR–16 (n = 1600) (Suppliment Fig. 1S2); 1821 DEGs targeted by 9 DEMs amongst which 7 upregulated DEMs including miR–491 (n = 92), miR–24 (n = 98), miR–650 (n = 341), miR–523 (n = 4), miR–422 (n = 99), miR–142 (n = 409), miR–1973 (n = 10) and 2 downregulated DEMs including miR–519 (n = 95), miR–665 (n = 673) identified in the IBD group (Suppliment Fig. 1S3). The UC and CD groups had 149 DEGs in common, while 110 DEGs fitted all the three groups (Fig. 1d), implying overlapping genetic constituent within CD, UC and IBD. The PPI network was visualized using Cytoscape among 571, 300, 947, 762 proteins in the CD, UC, upregulated and downregulated IBD, groups respectively, followed by analysis of the most connected nodes, considered as key regulators of pathways and biological functions (Suppliment Fig. 2S). Among the eleven topological analysis methods in CytoHubba, the MCC method, which was based on the degree of connectivity of each protein and the size of the interactions connected to the rest of the network, was proposed to have superior accuracy of identifying hub genes encoding proteins in the network [17]. Hub genes, being central element of the PPI network, serve as prospective biomarkers, therapeutic targets and novel tool for analyzing crucial mechanisms regulating disease processes.
The top ranking hub gene of the CD group was SOCS3 (Suppressor Of Cytokine Signaling 3) upregulated by miR–650 (log2FC = 1.745, P value = 0.018561). Cheng et al reported the role of miR–19b in inhibiting inflammatory response by downregulateing SOCS3 to alter chemokine production in CD [25]. PPI from cytoscape showed cytoplasmic expression of SOCS3 in most tissues and additionally in the plasma membrane; among blood cell types, it was specific to neutrophils. SOCS3 was expressed in the cells of epithelium and lamina propria in the colon in IBD, UC and CD cases [26]. SOCS3 as a member of the SSI (STAT-induced STAT inhibitor) family act to inhibit cytokine signal transduction through the JAK/STAT pathway [27]. SOCS3 also acted as a substrate recognition component of a SCF-like ECS (Elongin B/C-Cul2/Cul5-SOCS-box protein) E3 ubiquitin ligase complex to mediate ubiquitination and subsequent proteasomal degradation of cytokine receptors in inflammation [28].
In the IBD group, the key gene SKP2 (S-Phase Kinase Associated Protein 2) was upregulated by miR–142–3p (log2FC = 1.145, P value = 0.029848), while ASB1 (Ankyrin repeat and SOCS box protein 1) was downregulated by miR–665 (log2FC = –1.03, P value = 0.009245). Zhang et al demonstrated the role of miR–665 in promoting apoptosis and colitis in IBD by inhibiting ER stress components XBP1 and ORMDL3 [29]. SKP2 was tissue enhanced in placenta (https://www.proteinatlas.org) with nuclear and cytoplasmic expression and low blood cell type specificity (PPI data); ASB1 was localized in the nucleoplasm (https://www.proteinatlas.org), had low tissue specificity, and the protein expression was mainly in the CNS, adrenal medulla, parathyroid gland and enteroendocrine cells (https://www.proteinatlas.org).
Among the top hub genes of each group, the top 7 upregulated hub genes of the CD group namely, SOCS3, WSB1, ASB6, UBE2D2, UBE2D3, CDC27, UBE2B were also expressed as the upregulated hub genes of the IBD group, while the WSB1 gene from the CD group was concomitantly present in the downregulated hub genes of the IBD group (Suppliment Fig. 2S). The DEGs in both CD and IBD groups were significantly enriched (PPI enrichment 1.0E–16) in pathways such as class I MHC mediated antigen processing related to ubiquitination & proteasome degradation (Fig. 3). Analysis of biological processes showed that the DEGs in both CD and IBD groups were significantly enriched in protein ubiquitination (Fig. 3). Intracellular antigens and cytokines induce proteasome, immunoproteasome containing PA 28 and other accessory particles, for further degradation of the ubiquitinated substrates by an ATP-dependent mechanism (Fig. 4a). Peptides in association with HSP chaperones are translocated by TAP to the endoplasmic reticulum (ER) lumen, where they are placed on the MHC class I complex, made of heavy chain and β2m (Fig. 4a). MHC I fold and assemble in ER, assisted by several ER chaperones. MHC class I complexes present peptides on the cell surface, for recognition by CD8+ T cells and NK cells. The SOCS3 and SKP2 genes, in the present study were up-regulated during the course of CD and IBD respectively concomitant with upregulation of pro-inflammatory cytokine TLR6 in both CD and IBD; upregulation of pro-inflammatory cytokines such as IL6, CCL22, IL–1A in IBD only, through activation of TLR- and TNF-signalling pathways via activation of NFKB (nuclear factor kappa B) pathway; JAK signaling pathway and cytokine-cytokine receptor interaction all leading to ubiquitin mediated proteolysis, by K63-linked polyubiquitin chains (Fig. 4b). The ubiquitin proteasome pathway, is essential for protein degradation carried out by three classes of enzymes E1, E2, and E3; E1 and E2 prepare ubiquitin chains that are then linked to proteins by the E3 [30]. The SOCS3, SKP2, and ASB1 are components of the SOCSbox in ECS complex of E3 ubiquitin ligase containing CUL5, RNF7/RBX2, Elongin BC complex and SOCSbox [28].
In TLR (toll-like receptor) mediated signaling (Fig. 4b), the microbial components like flagellin, peptidoglycan, and lipopolysaccharide are recognized by TLR6 (paralog of TLR2), which in association with TLR2, trigger the recruitment of the adaptor protein MyD88 (Myeloid Differentiation Primary Response 88), the TOLLIP (Toll Interacting Protein), and TIRAP (Toll/Interleukin–1 Receptor Domain-Containing Adapter Protein ), through interleukin–1 receptor–associated kinases IRAK1 and IRAK4, in combination with RNF152 (Ring Finger Protein 152/ RING-Type E3 Ubiquitin Transferase RNF152) to stimulate K63-linked polyubiquitination. This complex, in turn, creates a binding platform for the assembly of the signaling genes, the TAK1, TAB1, TAB2, the TGF-β-activated kinase 1, TAK1-binding protein 1, 2 respectively, to cause the attachment of linear polyubiquitin chains on NFKB (Nuclear Factor Kappa B) essential modifier (NEMO) from the inhibitor of Kappa B kinase (IKBK) complex comprising CHUK (Conserved Helix-Loop-Helix Ubiquitous Kinase), IKBK-β/IKBKB (Inhibitor Of Nuclear Factor Kappa B Kinase Subunit Beta), and NEMO. The NEMO from the IKBK complex mediate K63-linked polyubiquitin binding on RNF152 to facilitate IKBK-β activation and successive phosphorylation of inhibitor of Kappa B alpha (IKBA) for ubiquitination with K63 linkages by the E3 ligase ECS complex to induce IKB degradation. In ubiquitin-mediated JAK signaling (Fig. 4b), cytokine-induced receptor dimerization results in tyrosine phosphorylation of JAK2 followed by binding of SOCS protein at its SH2 domain leading to ECS complex mediated K63 polyubiquitination and proteasomal degradation of JAK2 and termination of JAK signaling process. In TNF sinalling pathway (Fig. 4b), tumor necrosis factor (TNF) binds to the tumor necrosis factor receptor (TNFR)1, to trigger the recruitment of TRADD (TNFR1-Associated Protein), TRAF2/5 (TNF Receptor Associated Factor 2/5), RIP1 [Receptor (TNFRSF)-Interacting Serine-Threonine Kinase 1]. This, in turn, causes binding of TAK1, TAB1 and TAB2 ultimately leading to NFKB signaling pathway followed by K63-linked ubiquitin mediated proteolysis of IKBA.
Among the genes screened from the differentially expressed genes in UC, CIRH1A (Cirrhosis, Autosomal Recessive 1A) gene, in the present study was downregulated by miR–16–5p (log2FC = –1.195, P value = 0.04479). CIRH1A showed low blood cell type and tissue specificity (PPI data) with nucleolar expression in several tissues (https://www.proteinatlas.org), mainly in neuronal cells. CIRH1A, encode WD40-repeat-containing protein, involved in nucleolar processing of small subunit pre–18S rRNA. In the GO and pathway analysis, the top enriched pathways in UC included rRNA processing and modification in the nucleus and cytosol (Fig. 3). The CIRH1A gene, specific to the UC group, was down regulated concomitant with downregulation of U3 snoRNP complex, 90S pre-ribosome components, pre–40S, pre–60S, and export factors components (Fig. 4c). Impairment of ribosome biogenesis induce ER stress leading to proinflammatory cytokine production via NFKB activation with IKK (IKBA kinase ) activity maintained at basal level via IRE1 (inositol-requiring ER-to-nucleus signal kinase 1), an ER stress sensor [31]. The significantly downregulated genes in UC, in the present study, included receptor families such as TLR6 via NFKB signaling pathway; downregulation of pro-inflammatory cytokines such as IL6, IL–1A, IFNG through activation of JAK signaling pathway and cytokine-cytokine receptor interaction (Fig. 4b). IL–6 enhanced upregulation of rRNA transcription stimulated the MDM2 (mouse double minute 2 homolog)-mediated p53 proteasomal digestion, by reducing the availability of ribosome proteins for MDM2 binding [32].
{kind=link}
{kind=link}
{kind=link}
{kind=link}