Helminths have co-evolved with humans to immunomodulate their hosts. This includes schistosomes, which affect at least 200 million people in Brazil, sub-Saharan Africa, the Middle East, and Asia (1). The main schistosomes affecting humans include Schistosoma mansoni, Schistosoma haematobium, and Schistosoma japonicum, with S. haematobium comprising the majority of infections. All three of these species secrete immunomodulatory proteins, such as omega-1 (27), kappa-5 (28), SJHME-1 (29), Sj16 (30), and the interleukin-4-inducing principle of Schistosoma mansoni eggs (IPSE) (9), also called α-1(10). These immunomodulatory products of schistosomes have been postulated to exert far-reaching regulatory effects on host responses to not only schistosomiasis, but also co-infections.
Schistosomiasis is co-endemic with many other pathogens, including Plasmodium (31), HIV (31), mycobacteria (32), other helminths and parasites (33), and also Enterobacteriaceae such as Salmonella (34). Another member of Enterobacteriaceae that may co-infect large numbers of patients with schistosomiasis is E. coli.
Of the various types of disease-causing E. coli, uropathogenic strains are a common cause of infection in male and females, ranging from neonates to the elderly. One-quarter of all girls and women will experience multiple bacterial urinary tract infections (UTI) (35). Three different publications have linked urogenital schistosomiasis with bacterial UTI (3–5). However, two other papers have debated whether there is a causal connection between these infections (6, 7). There are multiple hypotheses that could explain a potential cause-and-effect relationship between S. haematobium infection, the primary cause of urogenital schistosomiasis, and UTI. It is possible, for instance, that detection bias contributes to increased reporting of UTI among patients with urogenital schistosomiasis. In this scenario, increased urine testing and urinary symptom tracking of individuals living in endemic areas leads to incidental discovery of UTI. Another potential hypothesis explaining a causal link between urogenital schistosomiasis and UTI is related to urinary tract obstruction. Schistosome egg deposition in the bladder initiates granulomatous fibrosis. In severe cases, this fibrosis can obstruct the urinary tract, contributing to urine stasis and bacterial overgrowth. Finally, it is possible that S. haematobium infection leads to immune skewing that hinders host clearance of bacteriuria. In support of this theory, we showed that S. haematobium eggs injected into the mouse bladder wall renders BALB/c mice, which are typically UTI-resistant, highly sensitive to subsequent UTI (8). The ability of S. haematobium eggs to enhance susceptibility to urinary tract co-infection relies upon IL-4, and also depended upon this cytokine dampening NK T cell activity. These findings lend credence to the possibility that the increased sensitivity of patients with urogenital schistosomiasis to UTI is orchestrated by schistosome-based immune modulation.
The egg-associated mediators which coordinate this IL-4-dependent phenomenon may include IPSE. This molecule is secreted by schistosome eggs in large amounts (11), and exhibits numerous immunomodulatory features. IPSE received its moniker because it ligates IgE- Fcε receptor complexes on the surface of basophils and mast cells and thereby induces IL-4 secretion (12–14). In a presumably similar manner, it activates B regulatory cells (Bregs) (15). The S. mansoni ortholog of IPSE called S. mansoni chemokine-binding protein (smCKBP) can sequester a number of chemokines (16). In addition, IPSE contains a nuclear localization sequence which traffics the protein to host cell nuclei (17, 18), where it alters transcription (19, 20).
Based on IPSE’s capacity to trigger IL-4 secretion and modulate host immunity more broadly, we posited that mouse exposure to S. haematobium-derived IPSE would be sufficient to augment UTI sensitivity. We sought to examine this postulate for two reasons. Firstly, we wished to delineate IPSE’s immune mechanisms given its association with urogenital schistosomiasis. We also sought to ascertain whether and how IPSE would be efficacious against detrimental, UTI-associated bladder inflammation. This goal was based on our past observations that a single injection of IPSE prevents severe bladder inflammation triggered by ifosfamide exposure (12, 20, 21).
Taking into account IPSE’s profound effects on other types of bladder inflammation, we were surprised by our finding that IPSE did not alter clearance of subsequent UTI. Specifically, IPSE did not change mean bacterial cfu over a week of daily urine sampling. IPSE also did not modify the distribution of bacterial cfu; in the mouse UTI model, sometimes mice in the same treatment group can cluster into bimodal distributions in terms of magnitude of bacteriuria. Monitoring levels of bacteriuria serially over multiple time points is important considering that the temporal kinetics of the mouse UTI model are highly dynamic. Our findings indicate that despite IPSE’s multi-faceted effects on host immunity, this molecule neither helps nor hinders clearance off bacteriuria.
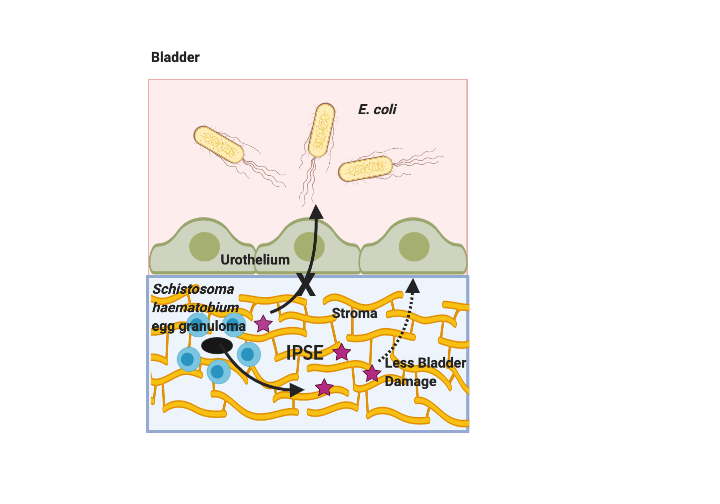
Although IPSE did not affect bacteriuria, it did alter UTI-induced bladder pathology. Namely, IPSE decreased bladder inflammation and edema, but did not ameliorate bladder contraction or urothelial denudation. By comparison, we have previously shown that H06-IPSE, the same ortholog tested in this study, reduces ifosfamide-induced bladder hemorrhage and histologically evident edema, but not bladder wet weights (12). In another publication we observed that H03-IPSE, another major S. haematobium ortholog, dampens ifosfamide-induced bladder wet weight increases (21). We did not measure bladder hemorrhage and wet weights in this study because bacterial UTI does not significantly alter these parameters (data not shown). Certainly, IPSE’s alleviation of UTI-triggered bladder inflammation is consistent with its known immunomodulatory properties.
After observing IPSE’s effects on UTI-associated bladder inflammation, we measured this molecule’s impact on bladder expression of anti-microbial peptides, an important set of immune responses to infection that evolved in early eukaryotes. Of the eight anti-microbial peptides examined, gene expression of lipocalin, beta-defensin, and alpha-defensin-4 were decreased by IPSE exposure. Lipocalin expression is associated with urinary tract infection (36). Transfection of the bladder urothelium with beta-defensin results in less mucosal damage, edema, and inflammation following UTI induction (37). These observations may appear puzzling when considered in the context of our findings; IPSE-mediated decreases in beta-defensin would thus be thought to enhance bladder damage. However, the functions of anti-microbial peptides are highly redundant, as a number of single anti-microbial peptide gene knockout mice, including beta-defensin-1 (38), have not demonstrated impressive UTI-associated phenotypes (39). Moreover, inhibition of certain anti-microbial peptides in the setting of UTI has been associated with less bladder damage (40). Thus, the effects of anti-microbial peptides on the bladder are complex. Finally, to our knowledge, alpha-defensin-4 does not have a critical role in UTI-associated immune responses.
Besides effects on anti-microbial peptide gene expression, IPSE has influence over basophil and mast cell immune responses. This molecule binds to Fcε receptor-associated IgE on the surface of basophils and mast cells (12–14). Ligation of these complexes could trigger histamine release and anaphylaxis, which are thought to have evolved as part of immune responses to parasites and their products (reviewed by Huang et al. (41)). Understanding IPSE’s ability to trigger histamine release is important to both comprehension of the pathobiology of schistosomiasis and defining this molecule’s safety profile as a potential therapeutic. When IPSE was intravenously administered to mice, systemic histamine levels were not elevated at either 5 or 30 minutes post injection. The 5-minute time point is particularly noteworthy considering that histamine release through anaphylactic degranulation is a very rapid physiologic process.
We also sought to characterize systemic aspects of host exposure to IPSE besides histamine release. In a repeated daily dose titration of intravenous IPSE at up to 2 mg/kg/day, we did not note any obvious adverse effects in terms of body weight trends, feed consumption, hematology, clinical chemistry, or gross pathology. There were very mild alterations in globulin levels and female liver weights (1 mg/kg/day of IPSE) that were not clearly significant. We observed sporadic, minimal changes in the kidneys, injection sites, and liver in some animals, but many of these changes were also seen in control animals.
Our findings have noteworthy caveats. For instance, the dosing of IPSE may not have been physiologic. Although IPSE is one of the most abundant egg-secreted proteins, it is difficult to estimate the exact tissue concentrations of IPSE that a S. haematobium-infected host would feature. Histamine responses, as well as sensitivity to histamine, are different in mice versus humans. Namely, humans are much more sensitive to histamine than mice. Furthermore, murine basophils have a tenfold lower histamine content (0.1 pg vs 1 pg/cell) compared with ther human counterparts (42). Although we attempted to perform a thorough assessment of any potential systemic pathology caused by IPSE exposure, it is possible that an important endpoint was not measured. In addition, differential binding affinities of IPSE for IgG and IgE could explain why there is no systemic anaphylactic degranulation. IPSE may bind to IgE with higher affinity than to IgG, but given there is several thousand fold excess of IgG in blood, IPSE may be “saturated” with IgG and cannot engage IgE on basophils.
In conclusion, IPSE is not sufficient to induce increased susceptibility to bacterial urinary tract co-infection. This suggests that IPSE is either not involved in potential enhanced vulnerability of patients with urogenital schistosomiasis to bacterial urinary tract co-infection, or that it works in concert with additional immunomodulatory factors produced by S. haematobium. IPSE’s apparent lack of toxicity and ability to alleviate UTI-triggered bladder inflammation and edema indicates that it may be suitable for continued development as a potential therapeutic for bladder inflammatory disorders.
{kind=link}