DENV infection disrupts TGF-β/Smad signaling by inhibiting phosphorylation and degrading Smad proteins
Smad2 and Smad3 are important members of receptor-activated Smads. Upon activation of TGF-β signaling, Smad2/3 get phosphorylated and translocated into nucleus and regulate TGF-β inducible transcription. To understand the association of DENV infection and TGF-β/Smad signaling, we utilized cell culture system of DENV replication with Huh-7 and HEK-293T cells. These cells are well documented with DENV replication.
Huh-7 cells were infected with DENV-2 strain NGC and viral replication was observed by specific qPCR at 24 and 48 hr post infection (hpi) (Fig. 1a). To check the phosphorylation status of Smad2 and Smad3, Huh-7 cells were infected with DENV followed by TGF-β treatment or left un-treated. TGF-β treated control cells were observed with higher expression of phosphorylated Smad2 and Smad3. A significant reduction was noted in the expression of phospho-Smad3 and no effect was observed on phosphorylation of Smad2 in DENV infection (p<0.05, Fig.1b and 1c). However, un-treated cells were noted with low expression of phospho-Smad2/phospho-Smad3 in controls and only difference was observed with phospho-Smad3. Therefore, it appears that DENV-2 infection may reduce the phosphorylation of Smad3 which leads to possible disruption of TGF-β signaling.
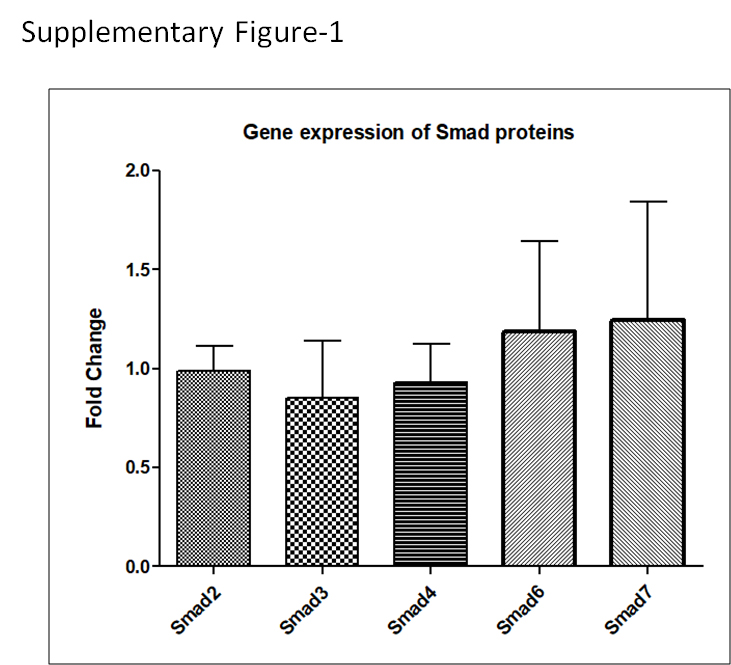
Further we investigated whether DENV infection also disrupts the non-phosphorylation status of Smad protein. To achieve this, we used HA or flag tagged constructs of Smad2 or Smad3 in HEK-293T cells. HEK-293T cells were transfected with constructs of Smad2 and Smad3 and 24h after transfection, these cells were infected with DENV and maintained till 48 hours post infection (hpi). Western blot analysis revealed expression of Smad2 and Smad3 was significantly disrupted by DENV infection at 48 hpi when compared with mock infected cells (p<0.05 and p<0.01; Fig. 1d and 1e). Smad signaling and regulation requires other assessor proteins such as Smad4 (co-smad), Smad6 and Smad7 (inhibitory Smads). We also utilized constructs of Smad4, Smad6 and Smad7 to see their expression pattern in HEK-293T cells with DENV infection. The western blot analysis of control and infected HEK-293T cells revealed significant reduction in the expression of Smad4, Smad6 and Smad7 at 48h post DENV infection (p<0.05; Fig. 1d and 1e). However, the gene expression analysis could not reflect any change in the Smad transcription level with DENV replication (Supplementary Fig.1). Overall, these results showed that DENV was clearly impairing the TGF-β signaling pathway.
Effect of Smads on DENV replication
To establish the effect of different Smads on DENV replication, we checked for viral load in the supernatants of DENV infected Smad overexpressing HEK-293T cells. qPCR data showed slight reduction in viral replication with Smad2, Smad3, Smad4 and Smad6 over expressing cells (p<0.05; Fig. 2a). Therefore we believe that these Smads may potentially reduce the viral replication.
Dengue virus NS1 Protein Inhibits TGF-β signalling by reducing the expression of Smads
DENV replication derailed the TGF-β/Smad signaling pathway by impairing the expression of Smad proteins. Several viral proteins have been known for their interfering role in the TGF-β/Smad signaling pathway [13,14,18]. We used DENV non-structural protein 1 (NS1), as it has been reported to interact with different host proteins [19] to find out whether NS1 has any role in the TGF-β/Smad pathway. To achieve the same, NS1 protein was co-expressed with other Smad proteins i.e. Smad2, Smad3, Smad4, Smad6 or Smad7, in HEK-293T cells. Except Smad7, we observed significant reduction in the expression of all Smads when co-expressed with NS1 (p<0.05 and p<0.01, Fig. 2b and 2c). Thus confirms the involvement of viral factor in TGF-β/Smad signaling.
Dengue virus NS1 blocks the nuclear translocation of Smad3
Nuclear translocation of Smad proteins is essential step of TGF-β/Smad signaling. As we confirmed earlier that NS1 reduced the expression of Smad2 and Smad3, so our next logical step was to examine whether or not NS1 alter nuclear translocation of these Smad proteins. Cells expressing Smad2 and Smad3 in the presence or absence of NS1 were fractionated into distinct cytoplasmic and nuclear fractions and analyzed by western blot. We have observed significant decrease in the expression of Smad3 in nucleus suggesting the possible inhibition of Smad3 nuclear translocation by NS1 (p<0.05; Fig. 3a and b).
E3 Ubiquitin ligase Smurf2 is required for regulation of Smads during DENV infection
Smad ubiquitination regulatory factor 2 (Smurf2) is an E3 ligase of HECT family. Smurf2 plays a key role in the editing of TGFβ/Smad signaling via ubiquitination of Smad proteins [9, 20]. Stub-1 or CHIP, another E3 ligase has been reported to promote ubiquitination of Smad proteins [10,21]. Here, we investigated the role of these E3 ligases, Smurf2 or CHIP during DENV infection.
NS1 and Smad protein constructs were co-transfected with Smurf2 or CHIP in HEK-293T cells. Western blot analysis revealed that Smurf2 has significant role in degradation of Smad3, Smad4, Smad6, Smad7 and had less effect on Smad2 (p<0.05; Fig. 4a and b). However, no difference was observed with CHIP co-transfection (Fig. 4c).
Smurf2 mutation reduce the effect of NS1 on Smad expression but not in CHIP-/- cells
We utilized construct of Smurf2 (C176A), which has mutation in the catalytic domain that make Smurf2 E3 liagse inactive [20,22-23] and also CHIP knockdown HEK-293T cells to see the effect on the expression of Smads with or without NS1. Smad4 forms a complex with Smad2 and Smad3 and translocates into the nucleus. So, we continued further experimentation only with these Smad proteins.
CHIP knockdown cells were prepared by using CRISPER-Cas9 method [24]. Smad2, Smad3, Smad4 and NS1 were co-expressed in the CHIP-/- HEK-293T cells. We observed higher expression of Smad3 and Smad4 in CHIP-/- cells and NS1 had less effect on their expression in these cells (Fig. 4d). Catalytically inactive Smurf2 had no effect with or without NS1 on the expression of Smad3 and Smad4, which confirmed the active involvement of Smurf2 during DENV infection (Fig. 4e).
Proteasome and lysosome mediated degradation of Smad3 and Smad4 by dengue virus NS1
E3 ligases marked their target proteins and promote their degradation via proteasome or lysosome mediated pathways. Host/cellular protein degradation is mediated by these two mechanisms [8]. Therefore, to understand the mechanism behind the escalated degradation of Smad3 and Smad4 by NS1, we utilized inhibitors of proteasome or lysosome mediated pathways, i.e., MG132 and chloroquine. Smad3, Smad4 were co-transfected with NS1 and transfected cells were treated with MG132 and chloroquine for 8 hrs prior harvesting and investigated levels of Smads by immunoblotting.
Following analysis of transiently transfected HEK-293T cell lysates by immunoblotting, we observed treatment with the proteasome inhibitor MG132 restores the expression of Smad3 compared to control or chloroquine-treated cells (Fig. 5a). However, we observed restoration in the expression of Smad4 with the addition of chloroquine but not with MG132, which clarified the mechanism of degradation of Smad4 was lysosomal mediated (Fig. 5a). Overall, this in vitro study suggesting NS1 exploits both of the proteasomal and lysosomal pathway to lower the expression of these Smad proteins.
NS1 interacts with Smad2, Smad3 and Smad4 complex
Smad2 and Smad3 form complex with Smad4 and translocate in to nucleus. To find out the physical interaction of Smad proteins and DENV NS1, we performed co-immunoprecipitation (co-IP) assay by using Smad proteins or NS1 as bait. Our co-IP data suggested that NS1 physically interacts with Smad proteins. We observed increase in degradation of these Smad proteins with NS1 when co-expressed with Smurf2. Therefore, we wanted to know whether Smurf2 is also interacting with NS1. Co-IP of NS1 and Smurf2 verified their interactions. Interaction of NS1 with Smad proteins and Smurf2 complex may inhibit the TGF-β/Smad signaling pathway (Fig. 5b).
{kind=link}