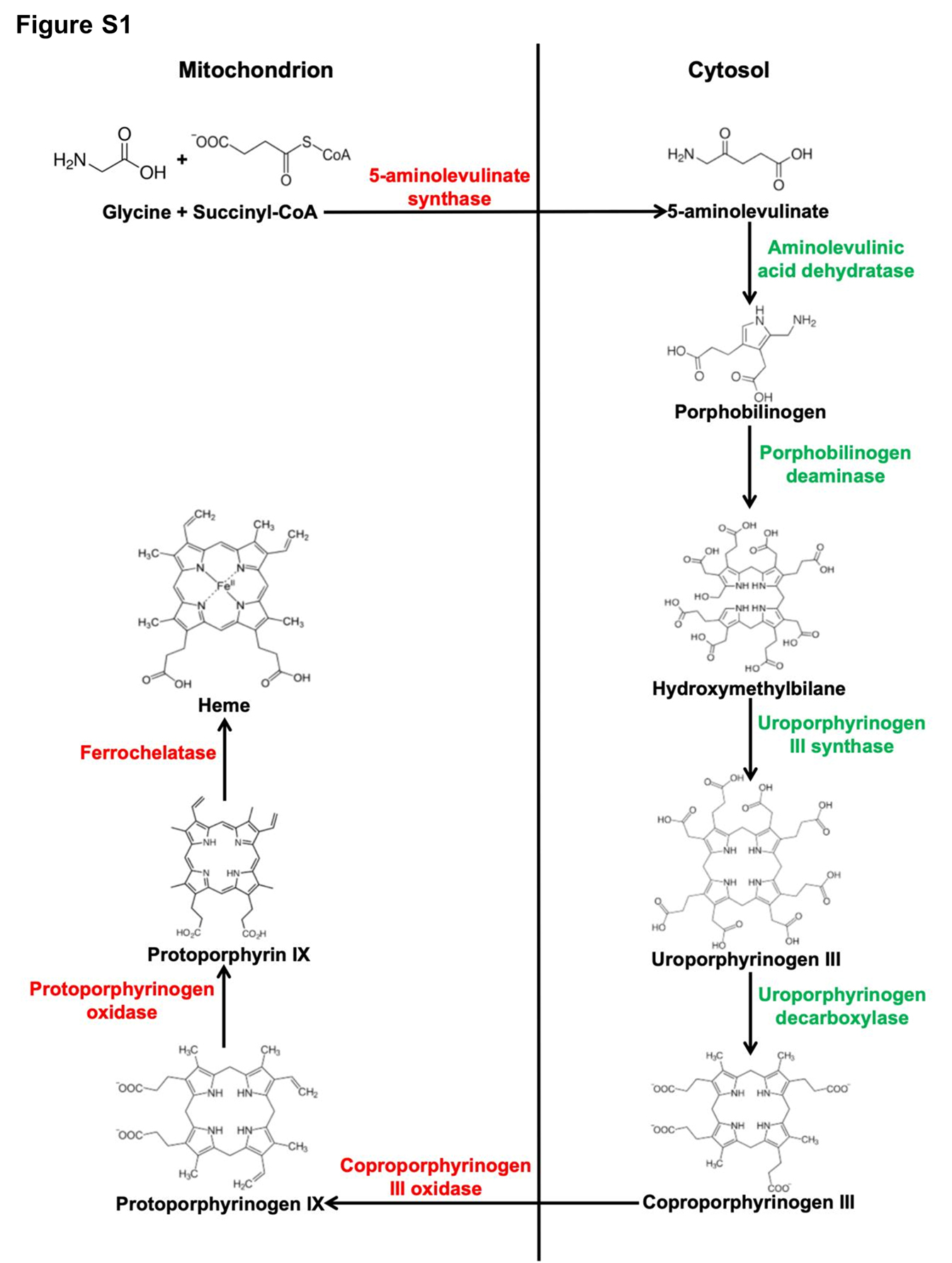
Human ALAD coordinates a previously unrecognized [Fe4S4] cluster
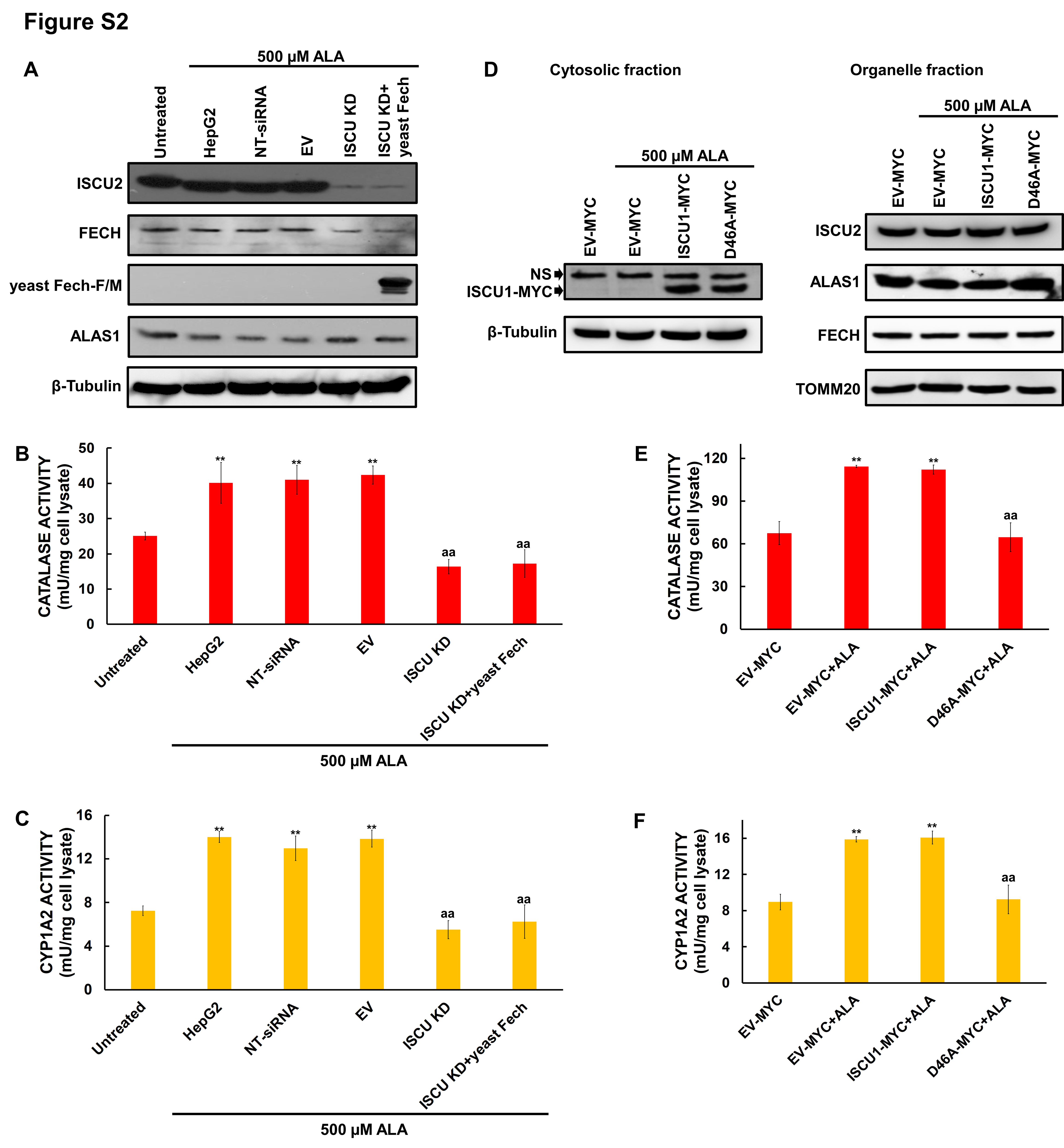
First, we explored whether there are as-yet-uncharacterized convergent points between the Fe-S and heme biosynthetic pathways. We first simultaneously knocked down the mitochondrial and cytosolic ISCU isoforms, which constitute the primary Fe-S biogenesis scaffolds in mammalian cells. Over-expression of yeast FECH, a non-Fe-S enzyme, did not restore heme biosynthesis in ISCU-deficient cells [Figures S2A-S2C and Supplemental Discussion (SD)]. Then we overexpressed a cytosolic ISCU mutant that impairs Fe-S biogenesis in the cytosol of mammalian cells and found that heme biosynthesis was inhibited (detailed in the SD and Figures S2D-S2F). Taken collectively, these results implied that one or more cytosolic heme biosynthetic steps was dependent on Fe-S biogenesis.
Then we screened the amino acid (AA) sequences of cytosolic heme biosynthesis enzymes for conserved LYR or LYR-like motifs to search for a potential Fe-S protein. Sequence alignment showed that human ALAD contains six cysteines, C119, C122, C124, C132, C162 and C223, and a LYR-like motif (Ala306-Phe307-Arg308) that is conserved among eukaryotic ALAD orthologs (Figure S3). Therefore, in terms of AA sequence and composition, ALAD appeared to represent a good candidate as an Fe-S protein.
As we have previously shown that de novo Fe-S biosynthesis occurs simultaneously in the mitochondrial and cytosolic compartments of mammalian cells23, we first determined whether human ALAD interacted with cytosolic HSC20. When the cytosolic fraction of HeLa cells was immunoprecipitated with anti-HSC20 antibody, the eluate contained ALAD (Figure 1A), suggesting that endogenous cytosolic HSC20 (C-HSC20) interacted with ALAD. Because HSC20 binds directly to LYR motifs, permitting transfer of nascent clusters from ISCU directly to recipient proteins16, 26, we then determined whether the AFR motif of human ALAD might be important for its interaction with HSC20. We transfected HeLa cells with plasmids expressing FLAG/MYC-tagged wild-type ALAD (ALAD WT) or AFR to alanine mutant (Mut AFR-AAA). Lysates from these cells were immunoprecipitated with anti-FLAG antibody, and the eluates were analyzed by western blotting. As shown in Figure 1B, wild-type ALAD interacted specifically with C-HSC20 but not with the lower-molecular-weight mitochondrial HSC20 (M-HSC20), as expected, given that ALAD is a cytosolic enzyme. No interaction between C-HSC20 and the AFR-AAA mutant was detected. Collectively, these data demonstrated that ALAD interacted with C-HSC20 and that the AFR motif of ALAD was important for the interaction.
To investigate whether human ALAD coordinates an ISC in human cells, we quantified 55Fe incorporation into C-terminally FLAG/MYC-tagged POLD1 (POLD1), a known [Fe4S4] protein that acquires its cluster in the cytosol 31, C-terminally FLAG/MYC-tagged ALAD WT and C-terminally FLAG/MYC-tagged Mut AFR-AAA in HEK293 cells transfected with either a pool of non-targeting siRNAs (NT) or with si-RNAs directed against human ISCU (si-ISCU). We found that ALAD and POLD1 bound comparable amounts of 55Fe in control HEK293 cells transfected with non-targeting siRNAs (Figure 1C and 1D). Mut AFR-AAA, which did not interact with C-HSC20 (Figure 1B), bound significantly less 55Fe than ALAD WT in NT HEK293 cells (Figure 1C, P<0.001). Compared with control cells, ALAD and POLD1 bound significantly lower amounts of 55Fe in HEK293 cells with defective ISC assembly machinery, achieved by silencing of human ISCU (Figure 1C and 1D, P<0.001). Taken together, these results demonstrated that ALAD binds iron, presumably in the form of an ISC, in human cells. To determine whether human ALAD can indeed coordinate an ISC in human cells, we then overexpressed C-terminally FLAG/MYC-tagged ALAD WT and Mut AFR-AAA in Expi293 cells and purified the recombinant proteins anaerobically. Anaerobically purified recombinant ALAD migrated as a single band on SDS-PAGE (Figure 1E) and exhibited a shoulder at ~420 nm in its UV-vis spectrum (Figure 1F), suggesting the presence of [Fe4S4] cluster(s)32, 33, 34.
We next expressed N-terminally MBP-tagged human ALAD (MBP-ALAD) in growing bacteria along with the A. vinelandii isc operon, which encodes the basic Fe-S biogenesis proteins and can assist iron-sulfur cluster assembly in bacteria, and purified the expressed protein anaerobically35, 36. The anaerobically purified MBP-ALAD was brown (Figure 1G), migrated as a single band on SDS-PAGE (Figure 1H), and also exhibited a shoulder at ~420 nm in its UV-vis spectrum (Figure 1I).
To validate the presence of [Fe4S4] cluster(s) and determine the stoichiometry of the [Fe4S4] cluster(s), a 57Fe-enriched MBP-ALAD sample was analyzed by Mössbauer and EPR spectroscopies. The 4.2-K Mössbauer spectrum collected in a 0.53 mT magnetic field applied parallel to the direction of γ radiation (Figure 1J, black vertical bars) was dominated by a quadrupole doublet with parameters typical of [Fe4S4]2+ clusters [isomer shift (δ) of 0.44 mm/s and quadrupole splitting parameter (DEQ) of 1.24 mm/s, 79% of total intensity, red line]17, 37. In addition, the spectrum revealed a small amount of another quadrupole doublet with typical [Fe2S2]2+ parameters (δ = 0.28 mm/s and DEQ = 0.50 mm/s, 11% of total intensity, blue line). Given that we measured that wild type ALAD binds 3.5 Fe per protein monomer (vide infra), the Mössbauer results imply that each ALAD monomer contains ~0.7 [Fe4S4]2+ and ~0.2 [Fe2S2]2+. [Fe2S2]2+ clusters are known to form as a result of [Fe4S4]2+ degradation35. Therefore, the minor quadrupole doublet corresponding to the [Fe2S2]2+ cluster could be a result of breakdown of a fraction of relatively unstable [Fe4S4]2+ clusters. Consistent with the Mössbauer analysis, the EPR spectrum showed no signal, ruling out the presence of Fe-S clusters with a half-integer spin ground state (i.e., [Fe2S2]+, [Fe4S4]+, [Fe4S4]3+, [Fe3S4]+) (Figure S4)17, 37. Taken together, these results demonstrate that each human ALAD monomer harbors one [Fe4S4] cluster when it is expressed along with the isc operon and purified anaerobically.
[Fe4S4]-ALAD has significantly higher enzymatic activity than zinc-ALAD
ALAD was previously identified as a zinc enzyme, and structures with zinc in the active site have been studied38, 39. Therefore, to determine whether the [Fe4S4] cluster of human ALAD is functionally significant, we compared the enzymatic activities of aerobically purified wild-type apo-ALAD (aero-apo-ALAD WT, see the Methods section for details), aerobically purified wild-type ALAD (aero-ALAD WT, aerobically purified wild-type ALAD prior to treating with EDTA, ), aerobically purified wild-type zinc bound-ALAD (aero-Zn-ALAD WT), and the anaerobically purified wild-type ALAD. As demonstrated by the ICP-MS analysis, aero-apo-ALAD WT did not contain iron or zinc, while aero-ALAD WT contained no zinc and a small amount of iron (Figure 2A). The enzymatic activity of aero-Zn-ALAD WT was approximately six times and two times higher than those of aero-apo-ALAD WT and aero-ALAD WT, respectively (Figure 2A and 2B, P<0.01). These results suggested that the enzymatic activity of apo-ALAD can be partly restored by zinc reconstitution. Wild-type ALAD expressed along with isc operon and purified anaerobically (ALAD WT) did not contain zinc (Figure 2A). Its enzymatic activity was approximately three times higher than that of aero-Zn-ALAD WT (Figure 2B the third bar versus the second bar, P<0.01). These results demonstrate that [Fe4S4]-ALAD has significantly higher enzymatic activity than zinc-ALAD.
The six highly conserved cysteines, Lysine 252, and the LYR motif of human ALAD are important for the enzymatic activity of human ALAD that contains a [Fe4S4] cluster in vitro
To identify the AAs important for the coordination/acquisition of the [Fe4S4], we mutagenized each of the six highly conserved cysteines, and Lys252, which has been shown to form a Schiff base with the carbonyl of one of the two ALA substrates in the enzymatic active site pocket (Figures S5 and S6). Additionally, we mutagenized the LYR-like motif of human ALAD to generate eight MBP-ALAD mutants (C119A, C122A, C124A, C132A, C162A, C223A, K252M, and Mut AFR-AAA). We expressed these mutants along with the bacterial isc operon and purified the over-expressed proteins anaerobically. To investigate whether the mutants coordinated a [Fe4S4] cluster, we plotted their molar extinction coefficients based on their UV-vis spectra. As shown in Figure 2C, none of UV-vis spectra of the mutants exhibited the absorption at ~420 nm, which is a hallmark feature of [Fe4S4] clusters. This observation indicates that these AAs are involved in either [Fe4S4] cluster coordination/acquisition or in maintaining the ALAD protein structure conducive to [Fe4S4] cluster coordination/acquisition. Importantly, the ALAD mutants all had significantly less iron and lower (or undetectable) enzymatic activities than ALAD WT (Figures 2A and 2B, P values <0.01). These results further confirm that the cysteines and LYR motif are important for [Fe4S4] cluster coordination/acquisition and the [Fe4S4] cluster is important for human ALAD enzymatic activity in vitro.
To determine whether the N-terminal MBP tag hindered the formation of a homo-octamer of ALAD subunits that constitute the previously defined active form of ALAD40, and to assess how [Fe4S4] cluster coordination affected ALAD quaternary structure, the anaerobically purified wild-type and mutant ALAD proteins were subjected to native gel electrophoresis. As shown in Figure 2D, most of the ALAD WT monomers formed ~640 kD homo-octamers, while all the ALAD mutants and the aero-apo-ALAD WT mostly assembled into ~480 kD homo-hexamers, a previously identified less active multimeric enzyme40. These results suggested that MBP-ALAD can form homo-octamers in vitro and that [Fe4S4] cluster coordination correlated with formation of higher proportions of ALAD homo-octamers.
The [Fe4S4] cluster of human ALAD is crucial for its enzymatic activity and heme biosynthesis in HepG2 cells
Next, we investigated the biological significance of the [Fe4S4] cluster of ALAD in human HepG2 cells. We co-transfected HepG2 cells with siRNAs to knockdown (KD) the expression of endogenous ALAD, and with a plasmid directing expression of either recombinant wild type ALAD (the ALAD WT group) or the ALAD mutant in which the AFR motif had been replaced by alanines (Mut AFR-AAA group). Effective KD of endogenous ALAD was achieved in HepG2 cells (Figure 3A). Recombinant ALAD WT or the Mut AFR-AAA mutant were well expressed with undetectable background expression of endogenous ALAD (Figures 3A and 3B, P values <0.01). We used ALAS1 protein levels and the activities of the heme-dependent enzymes cytochrome P450 1A2 (CYP1A2) and catalase, as readouts of the integrity of the heme biosynthetic pathway41, 42, 43. As shown in Figures 3A, 3D and 3E, there were no significant differences in the protein level of ALAS1 and catalase and CYP1A2 activities between the ALAD WT group and untransfected HepG2 cells. In cells expressing ALAD-Mut AFR-AAA, however, we observed significantly decreased ALAD, catalase and CYP1A2 activities in cell lysates as compared with untransfected HepG2 cells and with cells expressing ALAD WT (Figures 3A and 3C-3E, P values <0.01). These results demonstrated that the AFR-AAA mutant was non-functional and exhibited significantly lower enzymatic activity than wild-type ALAD. Therefore, overexpression of ALAD-Mut AFR-AAA did not restore heme biosynthesis that was suppressed by siRNA-induced ALAD deficiency. Given that the AFR motif of human ALAD is required for [Fe4S4] cluster acquisition, these results demonstrate that the [Fe4S4] of human ALAD is important for its enzymatic activity and heme biosynthesis in HepG2 cells.
Finally, we examined whether zinc addition affected ALAD enzymatic activity by adding zinc to the medium of mammalian cells over-expressing either WT ALAD or the Mut AFR-AAA (100 µM ZnCl2 for 48 hours). As shown in Figure 3A, levels of human metallothionein MT2A, whose expression is regulated by zinc44, 45, were greatly increased, suggesting that levels of cellular zinc indeed increased. However, there were no significant changes in either the protein levels of ALAS1 or the activities of ALAD, catalase or CYP1A2 after adding ZnCl2 to the medium (Figures 3A and 3C-3E). These results indicated that increasing intracellular zinc concentrations did not increase ALAD enzymatic activity in HepG2 cells overexpressing either wild-type ALAD or the AFR-AAA mutant. Notably, FECH protein levels were not affected by zinc addition, indicating that the coordination of [Fe2S2] cluster by human FECH was also unaffected by increased intracellular zinc levels (Figure 3A).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}