While ant queens retain the ancestral thorax morphology of flying Hymenoptera, we identified five major deviations of the worker thorax involving the exoskeleton, endoskeleton, neck muscles, leg muscles, and petiole muscles. Although our discussion focuses on Cataglyphis and Euponera, these modifications are present across all the ant workers examined (Table 1). Together, they represent the evolution of an enhanced power core for more effective foraging on six legs.
Fusion of the flexible articulated thorax into a rigid thoracic box
The thorax of a winged insect is essentially a flying engine with powerful wing muscles that vibrate articulated external plates called sclerites. As in other winged Hymenoptera, ant queens have huge indirect wing muscles (41% and 52% of thorax volume in Euponera and Cataglyphis, respectively) that fill the prominent mesothorax (T2) but also adjacent segments (Figure 1C and Figure 2). This is because the longitudinal muscles are attached to flexible cuticular invaginations (phragmata) of the mesonotum that extend into both the prothorax (T1) towards the front and the propodeum (IA) towards the back (Figure 2; the metathorax, T3, is much reduced dorsally and only serves for insertion of the hindwings [8]). Hence, much of the relatively small cavity of the prothorax of ant queens is occupied by wing muscles, with the neck muscles squeezed against the exoskeletal outer wall. Flapping of the wings is achieved not by direct muscle action, but by vibration of the entire thoracic box caused by the indirect wing muscles that deform the dorsal plates to which the wings are attached [21]. Flexibility of the thorax and free articulation of the dorsal plates (Figure 1A) are thus essential for flying, and winged ant queens conform to the general groundplan of Hymenoptera.
Ant workers lack wing muscles and the corresponding phragmata, and the plates of T2, T3 and IA are fused into a single rigid structure forming the posterior two-thirds of the thorax (Figure 1B, Additional file 1). The first third is the prothorax (T1) housing all the muscles that move the head, and it is greatly enlarged dorsally [10].
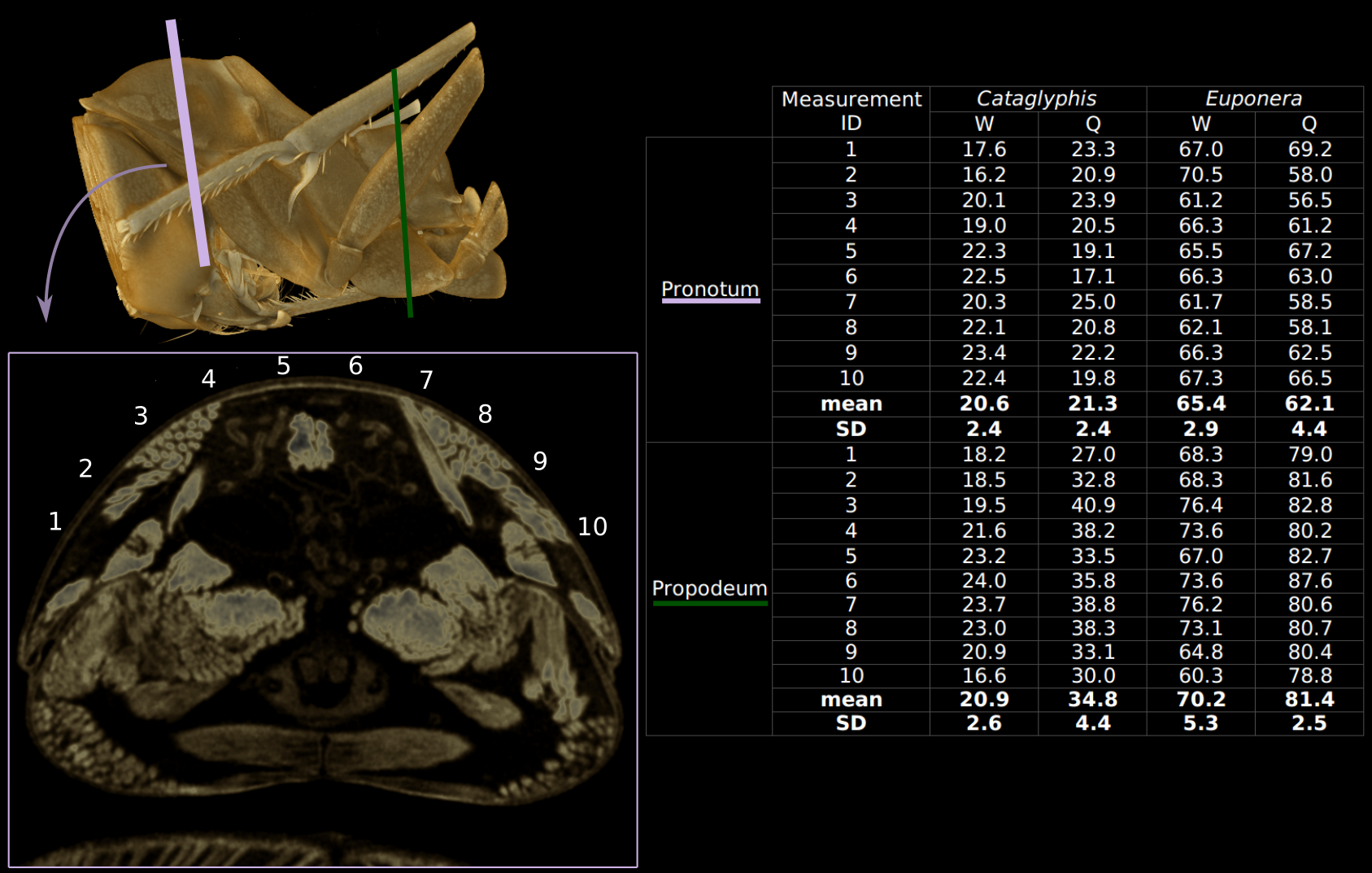
Thickness of Euponera cuticle is about three times that of Cataglyphis, in both queens and workers (Additional file 3). Thickness varies according to dorsal or lateral position of measurements within one segment, and variability is higher within one individual than between castes. Mean thickness is similar for workers and queens except for the propodeum of Cataglyphis.
Redesign of the endoskeleton for optimized bracing and muscle attachment
Inside the thoracic cavity, three invaginations of the sternae (ventral plates) rise up in front of each pair of legs, forming a row of rigid forked pillars. These furcae function as internal attachments for muscles that run between segments (head-to-prothorax, prothorax-to-mesothorax, metathorax-to-petiole), and muscles that move the legs (Figure 3). In queens, the main stem of the furcae rise not more than one-third of the thorax’s height before bifurcating towards the sides (Figure 4). This configuration results from the constraint of the huge longitudinal wing muscles that occupy the top two-thirds of the thoracic cavity (Figure 1C, Figure 2). In workers, the loss of wing muscles freed space within this cavity, allowing changes in size and shape of all three furcae.
The furcae of T2 and T3 are ancestrally fused into a single structure [18] (Figure 3). The T2 furca bears a narrow transverse bridge that connects the distal points of its forked arms (Additional file 4). In winged Hymenoptera, these lateral arms connect with the walls of the upper thorax via a short muscle, providing the necessary flexibility for flight. In ant workers this muscular connection is lost, and we found that the T2+3 furcal arms are fused with the walls of the thorax, forming an internal bracing structure that possibly increases the overall rigidity of the thoracic box.
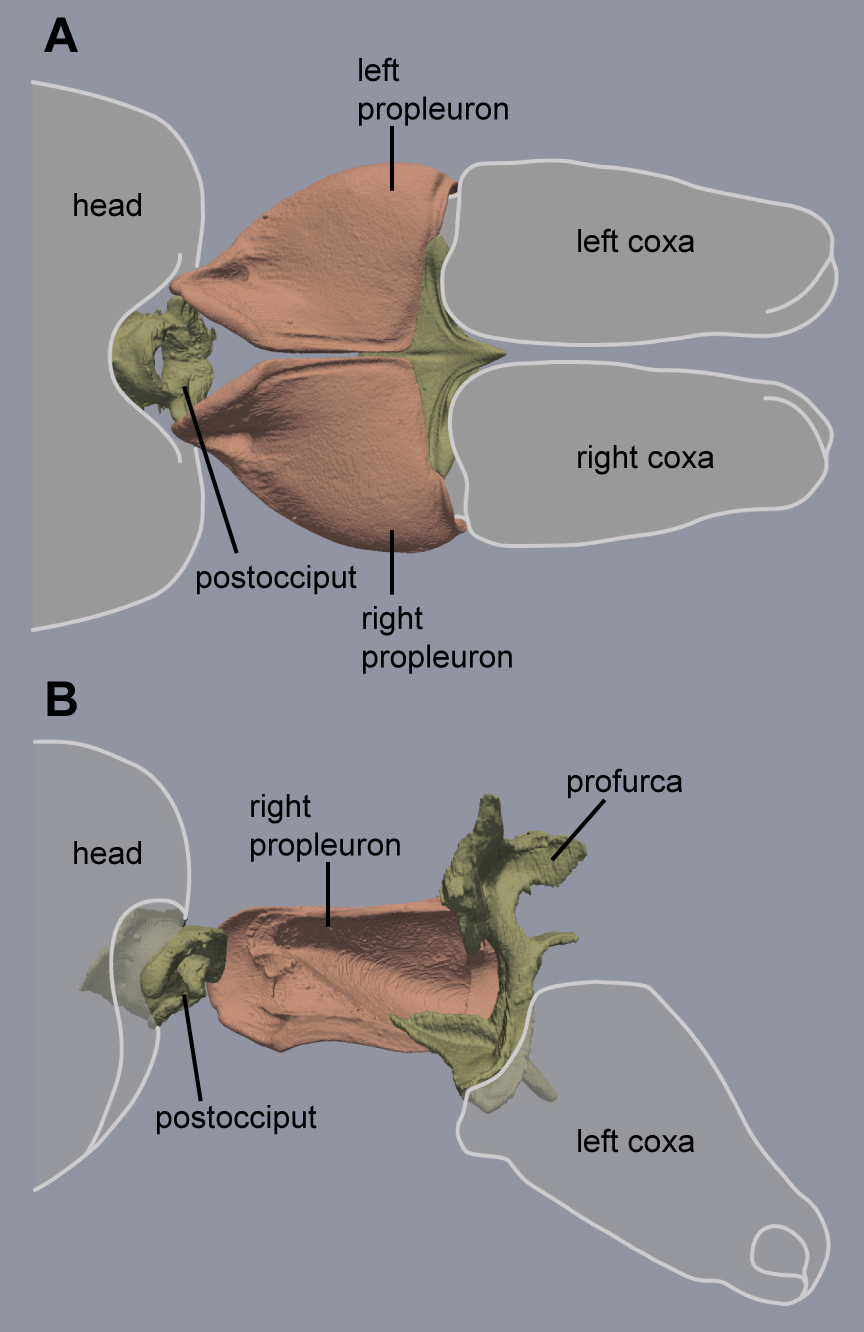
In contrast with the short and fused T2+3 furcae, the profurca (T1) is larger and more elaborate across Hymenoptera. Both the median stem and the forked lateral arms are stout and support a sizeable bridge, shaped like a thin platform (Additional file 5). This platform provides attachment surfaces for three neck muscle pairs on its anterodorsal side (muscles Idvm9, Ivlm1, and Ivlm3) and a key foreleg muscle on its posteroventral side (see below). In flying queens, this platform is horizontal to allow the longitudinal wing muscles and oesophagus to run dorsally (discussed in Keller et al. 2014 [10]). Consequently, the neck muscles attach to the platform at a low angle. However, the lack of wing muscles in ant workers removes this constraint, and the platform takes a more vertical orientation resulting in a greater attachment surface and more favourable angles of origin for the large muscles that insert directly on the back of the head and support it (see below).
Unlike the T2+3 furcae, the profurca is not fused with the thorax walls, instead its median stem articulates freely in front of the foreleg insertions while its lateral arms connect to the thorax walls with membranes and a short muscle [8]. The neck muscles act antagonistically to both foreleg muscles (see below) and the intersegmental muscles (Ivlm7) that connect the profurca with T2+3 furcae (Figure 3), transmitting stress from the head to all legs. Similarly, posture is maintained at the back of the thorax with intersegmental muscles (IIIvlm7) connecting T2+3 furcae to the petiole to transmit stress from the abdomen to the thoracic box.
Reorganization of the neck muscles
The neck articulation in Hymenoptera is composed of four skeletal elements (Additional file 6): the postocciput, a short cup-shaped extension of the back of the head; a pair of triangular propleura that are ventrally situated in T1; and the profurca which sits inside T1. The head’s postocciput has a strong articulation with the thorax via a stiff neck membrane [22] as well as the anterior apodemes of the propleura [18]. The propleura are never fused to one another and these large plates can shift antagonistically for sideways and rotational movement of the head [23].
No muscles inside the head are involved in the movement of the head relative to the thorax. Rather, all neck muscles are prothoracic, moving the head either directly by inserting on the postocciput or indirectly by inserting on the propleura. In both queens and workers, four pairs of direct muscles lift the head up-and-down (Figure 1C-D and Additional file 5). The lack of the anterior phragma in workers (associated with loss of wing muscles) affects especially muscle pair Idlm1. In queens, Idlm1 originates at the sides of their pliable phragma (Additional file 7), while in workers Idlm1 originates further back at the anterior margin of the roof of T2 (part of the rigid thoracic box; Figure 3). Consequently, the better anchoring of muscle Idlm1, its extra length and lower angle of insertion to the postocciput (Additional file 7) give greater support and strength to the worker head in comparison to that of winged queens. The direct muscles Itpm1 originate at the base of propleura, hence their geometry differs little between queens and workers, but Itpm1 is relatively larger in workers for both species (Figure 6). In contrast, muscle pairs Idvm9 and Ivlm3 are considerably affected by the queen-worker difference in the angle of the profurca platform (26° and 53° in Euponera queen and worker, respectively; 16° and 30° in Cataglyphis queen and worker; Additional file 5). In queens, as in wasps and bees, this platform is almost horizontal, and thus, these muscles have a very shallow angle to their origin. Contrary to queens, the upright inclination of the platform in workers allows a more favourable perpendicular angle of origin and a greater attachment surface for the equivalent muscles.
Two indirect muscle pairs are mostly responsible for sideways and rotational head movement by inserting on the anterior ends of the propleura. Ivlm1 originates on the profurca and thus benefits from the more favourable inclination of its platform in workers (Additional file 5). Idvm5 shows the most spectacular difference between queens and workers (Additional file 7). This large muscle pair originates at the pronotum (dorsal plate of T1) which is considerably larger across all worker ants. In addition to the increased area of attachment, the absence of wing muscles allows this muscle to support the propleura from a diagonal rather than a vertical angle. Both Idvm5 and Ivlm1 are relatively larger in workers for both species (Figure 6).
One crucial leg muscle differs between workers and flying queens
Back and forth movements of insect legs are controlled by a series of short muscles that originate within the thorax and insert around the opening of the coxae, the basalmost segment of each leg that articulates with the body. Leg flexion and extension, on the other hand, are controlled by muscles that reside entirely within the segments of the legs [18]. A notable exception is the external trochanter muscles. One per leg (Iscm6, IIscm6, IIIscm6), these muscles originate in the thorax but end in a long tendon that crosses the coxae and inserts on the trochanter (second leg segment) of all legs (Figures 4 and 5). Contraction of the external trochanter muscles causes depression of the leg, lifting the body and carrying the ant’s weight plus any load. Because these muscles are housed outside the legs, with only their tendons crossing the coxae, their action is independent from the rotational motion of the coxae, and force is transmitted efficiently from the thorax to the legs no matter if walking or standing.
While the muscles of the coxae and those internal to the legs do not differ much between the castes, our segmentations show that the external trochanter muscles are very distinct in length, geometry, and place of origin between queens and workers. In queens, the external trochanter muscles are shorter because they originate on the furcal arms of their respective segment, just below the longitudinal wing muscles (Figures 4 and 5). The absence of wing muscles in workers allows dorsal elongation of the mid and hind external trochanter muscles (IIscm6, IIIscm6), which attach either on the roof (Cataglyphis) or lateral walls (Euponera) of the rigid thoracic box (Figures 4 and 5, Additional file 8). Queen-worker differences are minor for the forelegs: in both castes external trochanter muscles Iscm6 originate at the lower surface of the T1 furcal platform (Figures 4 and 5). These muscles, therefore, act antagonistically to the neck muscles originating at the upper surface of this platform, helping to pull the head and transmitting force effectively from the head-neck articulation to the front legs. Workers in both species have a relatively larger IIscm6 than queens, and in Cataglyphis Iscm6 and IIIscm6 are also larger (Figure 6).
Enlargement of the petiole muscles
As with the head, all muscles responsible for articulating the free part of the abdomen are housed inside the thorax. Four muscle pairs (two dorsal, two ventral) insert on the anterior narrow end of the petiole, which protrudes in the back of the thorax. By acting antagonistically to each other, these muscles control both vertical and horizontal movements of the abdomen [18].
Our segmentations show that the two dorsal pairs differ strongly between queens and workers due to the presence/absence of the flight apparatus (Figures 4 and 5). Muscles IA1 and IA2 originate on the roof of the propodeum (middle and lateral, respectively), but because of the posterior end of the horizontal wing muscles in queens, these petiole muscles are squeezed in the narrow space between the posterior phragma and the exoskeleton (Figure 2). In workers, the lack of wing muscles means that muscles IA1 and IA2 fill most of the propodeal cavity (Figures 4 and 5). Although relative muscle volumes are similar between queens and workers (Figure 6), differences stand out in the fibre geometry in E. sikorae. First, IA1 has longer fibres in the worker than in the queen despite the smaller size of workers. In addition, IA1 fibres attach at significantly lower angles in the worker, resulting in greater efficiency (Figure 7). Second, IA2 fibres are longer in the queen and attachment angles are not significantly different between worker and queen. However, the worker has 30% more muscle fibres with a broad distribution of angles, revealing how much this muscle can fan out in the absence of wing muscles.
Marked differences between the two different subfamilies are seen in muscles IA1 and IA2 of workers. In Euponera workers, these petiole muscles originate on the entire roof of the elongated propodeum (Figure 4). In contrast, in the dome-shaped propodeum of Cataglyphis, the equivalent muscles do not extend forward and are relatively smaller in the worker than in the queen (Figure 5); instead the anterior roof is the origin for the external trochanter muscles (IIIscm6) of the hind legs (Additional file 8).
Comparison with other flightless insects
Flight is the ancestral state across the insect tree of life, and its evolutionary loss occurred multiple times independently [24]. While a broad survey of flightlessness across insects is beyond the scope of this paper, we considered whether the modifications described here are specific to social as opposed to solitary lifestyles. In general, the lack of massive wing muscles does not lead to the same modifications of the thorax in solitary taxa. A female mutillid wasp shows strikingly enlarged abdomen muscles (there is no petiole), attached to most of the dorsal roof and reaching the anterior mesonotum (Figure 8). This is congruent with the origin of the external trochanter of midlegs on T2 furca, instead of the propodeum roof. Given that mutillids are parasitoids that search for hosts underground and oviposit on the spot [25], this difference in thorax rearrangement likely reflects the absence of any selective pressures for prey transport. Indeed, added strength is not necessarily adaptive in solitary insects that are not central place foragers. In flightless moths (Lepidoptera) [26], ovaries reach into the posterior thorax, and the posterior phragma is retained to shield the ovaries; head muscles are not affected compared to flying males. Beetles (Coleoptera) fly with their hind wings only, hence flight musculature is confined to their enlarged T3 and does not conflict with the neck articulation. A functional approach makes it possible to interpret the internal adaptations associated with wing loss across insects showing divergent ground plans.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}