Microsatellite marker isolation and assessment
In the present study, a total of 44 pairs of microsatellite markers were isolated from the whole genome of Ae. albopictus, 11 pairs of which were highly polymorphic and were therefore chosen for microsatellite genotyping analysis (Table 1). The allele number of each locus ranged from 10 to 33, with a mean of 17.545 alleles per locus. The PIC values of each locus ranged from 0.334 to 0.925, with a mean value of 0.713. According to the definition of PIC values by Allah et al. [57], nearly all the markers selected were highly informative (PIC value > 0.5); the exception was BW-P18, which had a value of 0.357 and was considered to be an informative marker. The Micro-Checker results suggested that null alleles were present at all loci, with frequencies ranging from 0.064 to 0.157, with an average of 0.078 (Additional file 8: Table S5); however, frequency values smaller than 0.2 are considered to have no significant effect on the accuracy of data analysis by many studies [59, 65]. The linkage disequilibrium (LD) test showed that 302 pairs of loci of a total of 1870 (16.15%) across all locations were in significant LD after Bonferroni correction, while no consistency was found among them (Additional File 5: Figure S1).
Genetic diversity and variation
According to the results of the microsatellite data analysis, the observed number of alleles (na) in each Ae. albopictus population was very high, and the mean na value in each climatic region ranged from 6.909 to 8.091 without a significant difference. In contrast, the effective number of alleles (ne) ranged from 3.501 to 4.525, and the ne value increased from the temperate region (3.876) to the tropical region (4.144). The mean value of observed heterozygosity (Ho) for all the climatic regions was ca. 0.557, which was significantly lower than the expected heterozygosity (He) (ca. 0.684). The FIS value of each climatic region ranged from 0.266 to 0.359, and the Ae. albopictus populations significantly departed from HWE, except in two locations, SHJD and KZXZ (subtropical region, Table 2). Heterozygosity tests of all 17 Ae. albopictus populations based on the stepwise mutation model (SMM) revealed that nearly all the populations from the temperate and subtropical regions displayed significant population expansion with p-values < 0.05, while no significance was observed among all the tests for the Ae. albopictus populations from the tropical region after Bonferroni correction (Additional file 2: Table S2).
In all, a total of 25 coxI haplotypes were observed among 497 Ae. albopictus individuals (GenBank ID: MN651301-MN651325) based on an analysis of the coxI sequences. The haplotype indices (i.e. Hd, H, and π) changed dramatically across Ae. albopictus populations from different climatic regions (Additional file 3: Table S3). The haplotype diversity (Hd) ranged from 0.074 (ZGND, temperate region) to 0.750 (JYJB, tropical region), with the nucleotide diversity (π) ranging from 0.014 (BJLG, temperate region) to 0.190 (JYJB, tropical region). The average number of nucleotide differences (k) ranged from 0.067 (BJLG, temperate region) to 0.929 (JYJB, tropical region), and the number of polymorphic sites ranged from 1 to 5 across all the populations. Overall, the tropical Ae. albopictus populations showed the highest diversity, with mean values of Hd, π, and k reaching 0.657, 0.156, and 0.763, respectively, whereas the temperate populations showed the lowest diversity.
Population structure and differentiation based on microsatellite analysis
In the present study, according to the microsatellite analysis results, all the Ae. albopictus populations were adequately allocated to two clades with significant genetic differences, and the best K value, as determined via the ΔK method of Evanno et al., was also equal to two (Fig. 2a). Combined with the STRUCTURE bar plot analysis, the Bayesian clustering analysis showed that all the Ae. albopictus populations were adequately allocated to two clades with certain locations from the subtropical and temperate regions genetically isolated from the other locations. (Fig. 2b&c). Moreover, a total of 86.4% of the variation was explained by 50 PCs in the discriminant analysis of principal components (DAPC) analysis; the results revealed two genetically isolated groups, and there was no clear relationship between the Ae. albopictus population structure and its distribution across climatic regions (Fig.2d).
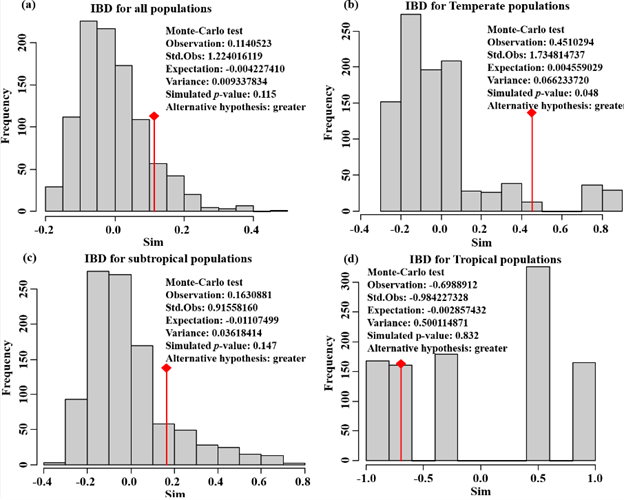
The AMOVA results are presented in Table 3. Of the total genetic variation partitioned, 31.40% could be attributed to differences among individuals within populations and 63.04% to differences within individuals (FIS = 0.33253, FIT = 0.36962, and all p < 0.00001). Meanwhile, the pairwise FST values between populations ranged from 0.008 (ZGND, temperate region and NNXD, subtropical region) to 0.141 (SHJD, subtropical region and JKCH, tropical region). Genetic differentiation was significant between all the sampled populations after Bonferroni correction (p < 0.05) except for five pairs of FST values among the subtropical populations NNXZ, NJTH, NJDX, KZXZ, and temperate populations ZGND, BHBG, QDDX, and HBSD (Additional file 4: Table S4). In contrast to the high individual variation, a slightly significant positive correlation was observed among only the temperate populations via isolation by distance (IBD) analysis (R2 = 0.6614, p = 0.048), while no such evidence was observed in the other two regions (Additional file 6: Figure S2).
Haplotype network and phylogenetic analysis based on coxI sequences
Three major haplotype clades, distributed across the three central climatic regions and closely related to each other, were reconstructed via the TCS network with 497 coxI sequences. Haplotype Ⅰ (H1) of Clade Ⅲ was the most frequent haplotype and was distributed from the tropical to the temperate region with an increasing trend. Nearly all the other haplotypes derived from H1 with one or two mutations, with Clade Ⅰ mainly distributed in the tropical region and Clade Ⅱ mainly distributed in the southern subtropical region (Fig. 3b).
A phylogenetic tree of all 25 haplotype sequences, combined with a heatmap analysis, demonstrated that the 25 haplotypes were divided into three major well-supported clades. As expected, Clade Ⅰ was separated from Clade Ⅱ and Clade Ⅲ with 100% bootstrap support and included haplotypes H9, H10, H11, H12, H19, and H20, which were observed in only the tropical region. In comparison, Clade Ⅱ diverged from Clade Ⅲ with lower bootstrap support (70.74%) and included five haplotypes (H2~H3 and H5~H7) that were distributed in only the southern subtropical region. Clade III was an admixture group containing all the remaining 14 haplotypes, two of which were observed in the tropical region and four in the climatic regions (Fig. 3a). Meanwhile, the Tajima’s D and Fu’s Fs values were negative, with no statistically significant p-values except in population JKCH (tropical region, Additional file 3: Table S3). Based on the coxI sequences, the mismatch analysis results showed that the Harpending raggedness indices for all three haplotype clades were relatively low (ranging from 0.1054 to 0.1812, p > 0.05), and unimodal mismatch distributions were observed among the different Ae. albopictus populations (Additional file 7: Figure S3).
Migration and correlation analyses between genetic indices and environmental factors
As illustrated in Fig. 4, all five genetic indices (i.e. ne, Shannon index, Hd, H, and π) and two environmental factors (i.e. temperature and rainfall) contributed equivalently to the first axis of the PCA, explaining up to 95.2% of the variation; the exception was the environmental factor latitude, which contributed more to the second axis, with a proportion of 17.5%. Combining these results with hierarchical clustering via MFA, all 17 Ae. albopictus populations were clustered into three groups, of which cluster II and cluster III were closely related to each other. When environmental factors were regarded as the major influencing factors, the molecular diversity indices (i.e. Hd, k, and π) of the populations in the tropical region were significantly higher than those in the other regions (Additional File 3: TableS3).
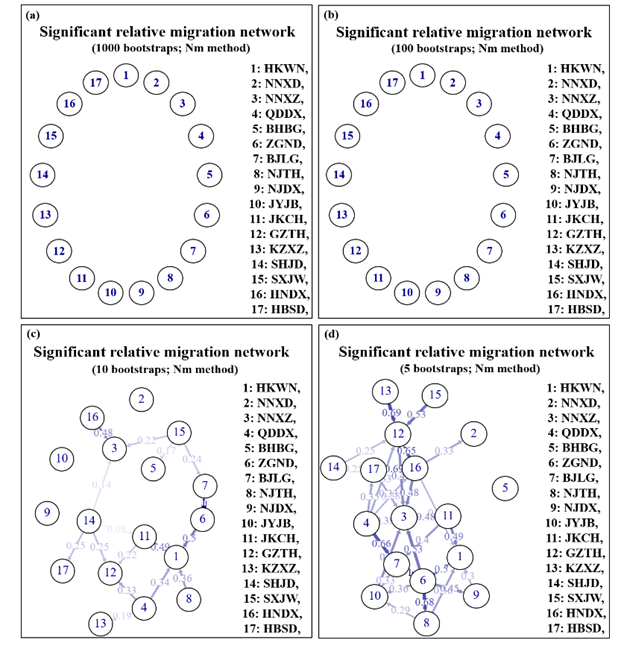
Migration patterns were assessed using divMigrate networks representing all 17 Ae. albopictus populations. As expected, Ae. albopictus was observed to have migrated frequently between the tropical and temperate areas in both directions (Fig. 1). A total of four major migration trends were observed among the different climatic regions with high gene flow (Nm > 0.4). Two trends were found to have originated from the tropical and subtropical areas and reached Hebei and Beijing in the temperate area, while another two arrived in Guangdong and Guangxi from the southern subtropical area. Compared with the south to north routes, the latter migration routes were substantially larger.
{kind=link}
{kind=link}
{kind=link}