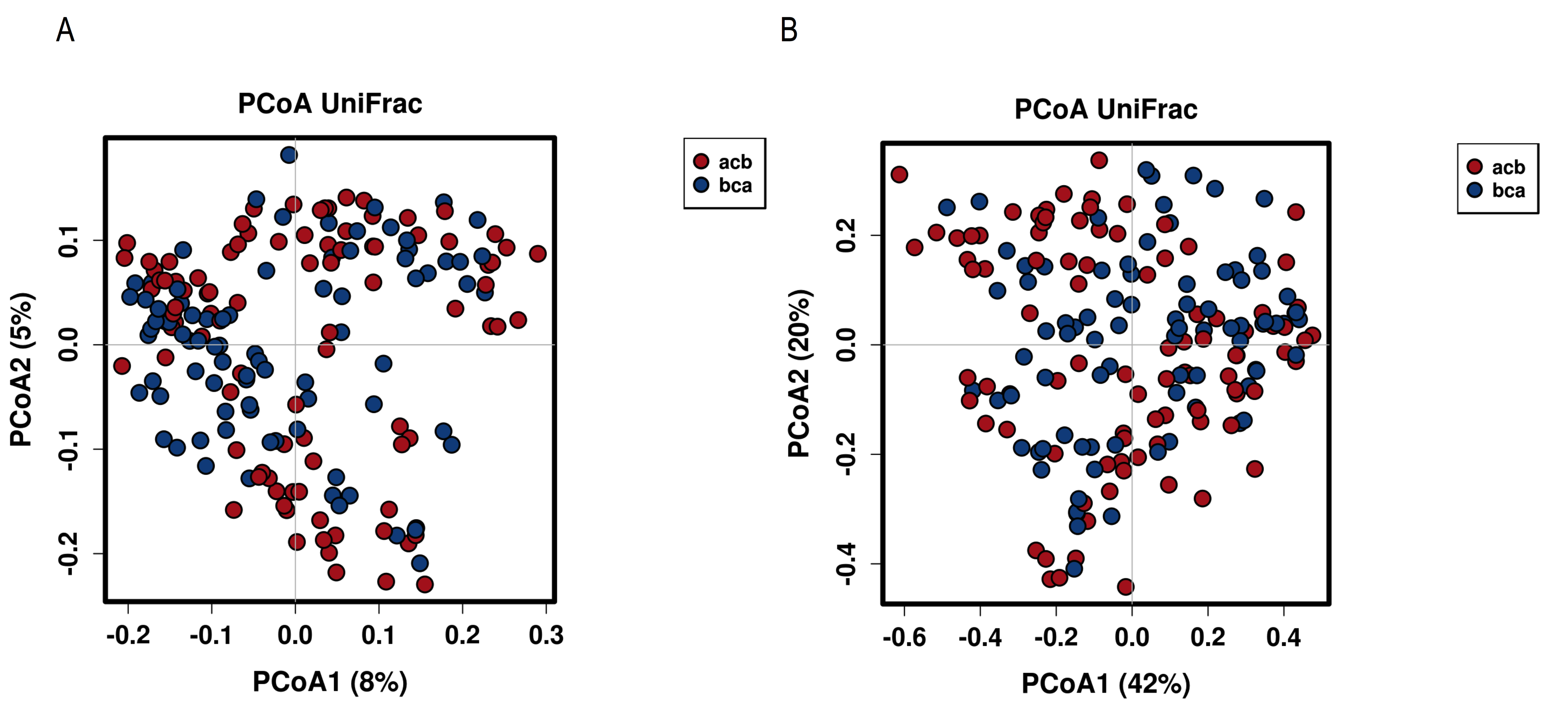
Several studies have been conducted on the effect of diet on the gut microbiota in humans and rodents, and more studies are emerging in dogs. However, the gut microbiota-diet relationship is complex and challenging to characterise as many factors may influence the outcome [2]. In our study, all dogs were receiving the same baseline diet, were the same breed, similar age, similar body condition and lived in the same environment; which served to eliminate many confounders factors that could influence the results. In general, we observed that the treatment diets had a significant residual impact on the faecal communities of all dogs and results were dependent on the composition of the gut microbiota at the start of the intervention.
Analysis of the alpha diversity between the different categories of diet, showed that the high-insoluble fibre diet and hydrolysed diet have a higher Shannon diversity in comparison with the raw meat, high-protein diet. However, when the analysis was done per subject, the difference in Shannon diversity was minimal between diets.
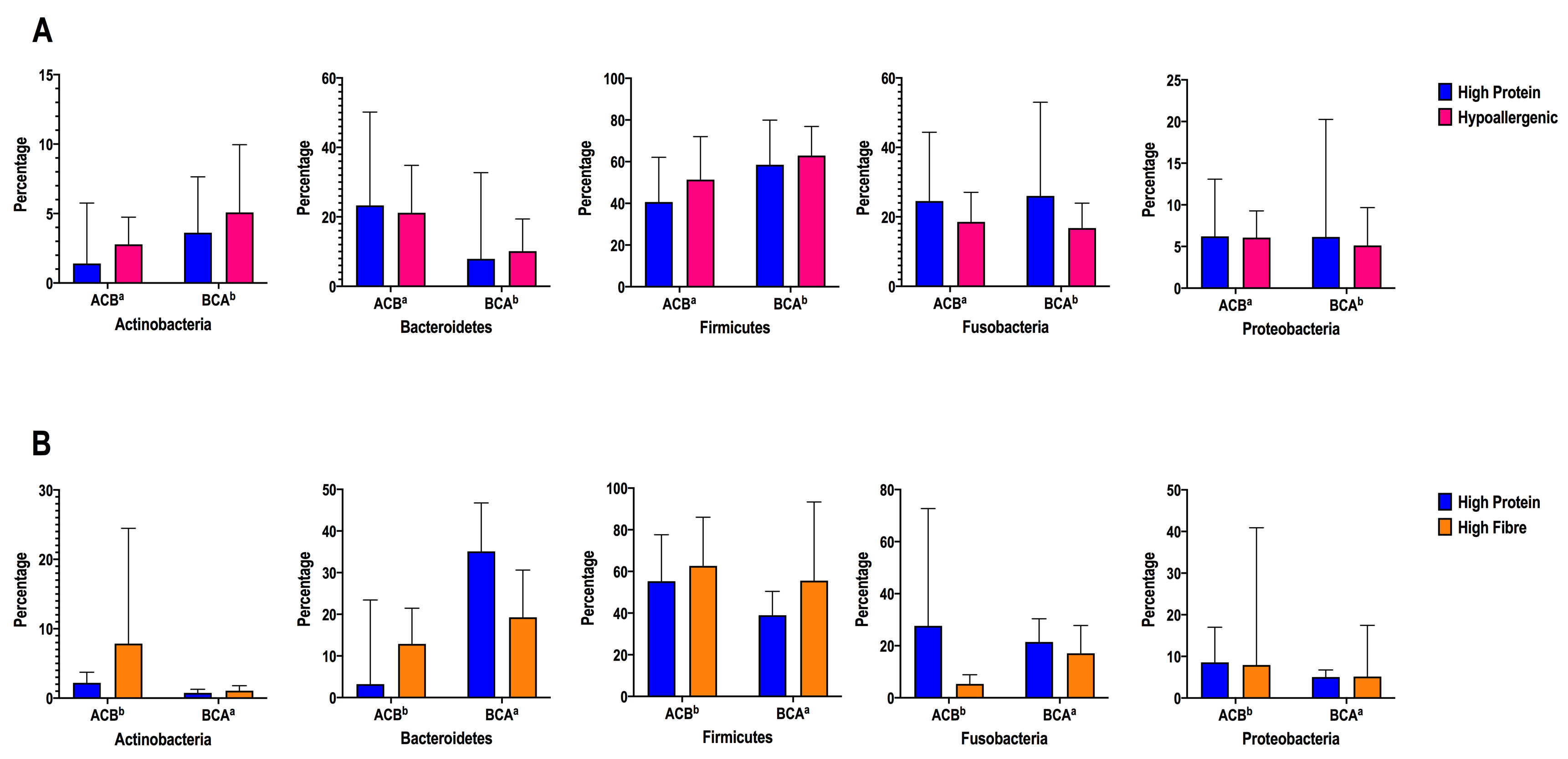
Studies investigating the direct impact of protein on gut microbiota composition and functionality have shown that protein quality and source are as important as total amounts in people, but less so in dogs [2,16,44]. Analysis of the gut microbiota showed that diet C (raw all-meat/high protein diet) in our study was characterised by an overrepresentation of bacteria belonging to the Fusobacteria phylum. This contrasts with a previous study made in obese and lean dogs with high-protein dietary intervention, where the changes in Fusobacteria were relatively small, although the levels of protein differed between studies (49.38% vs. 69% in ours) and was of a shorter duration [21]. Another study where the dry commercial diet was changed to minced beef, also showed minimal changes in the Fusobacteria content [18]. High levels of Fusobacterium sp. have been observed in carnivores of multiple species [3, 45, 46]; and in humans, increases in the levels of Fusobacterium are seen in people consuming a diet high in red and processed meats and are associated with an increased risk of development of colorectal cancer [47, 48]. In one study of dogs fed a red meat diet for 9 weeks, an increased Fusobacterium abundance (~15%) was present at 6 weeks, suggesting these changes may take time to develop (20). Diet C in our study consisted mainly of horse carcass, which is a vastly greater percentage of protein in diet than most commercial diets (prescription or supermarket brand). Digestibility of scrap meat may be lower than for high quality protein due to the high amount of connective tissue, and digestibility of macronutrients may also influence the colonic microbiome [49]. Additionally, most commercial dog food does not contain horse protein, which may be biologically different than other sources of protein [50, 51].
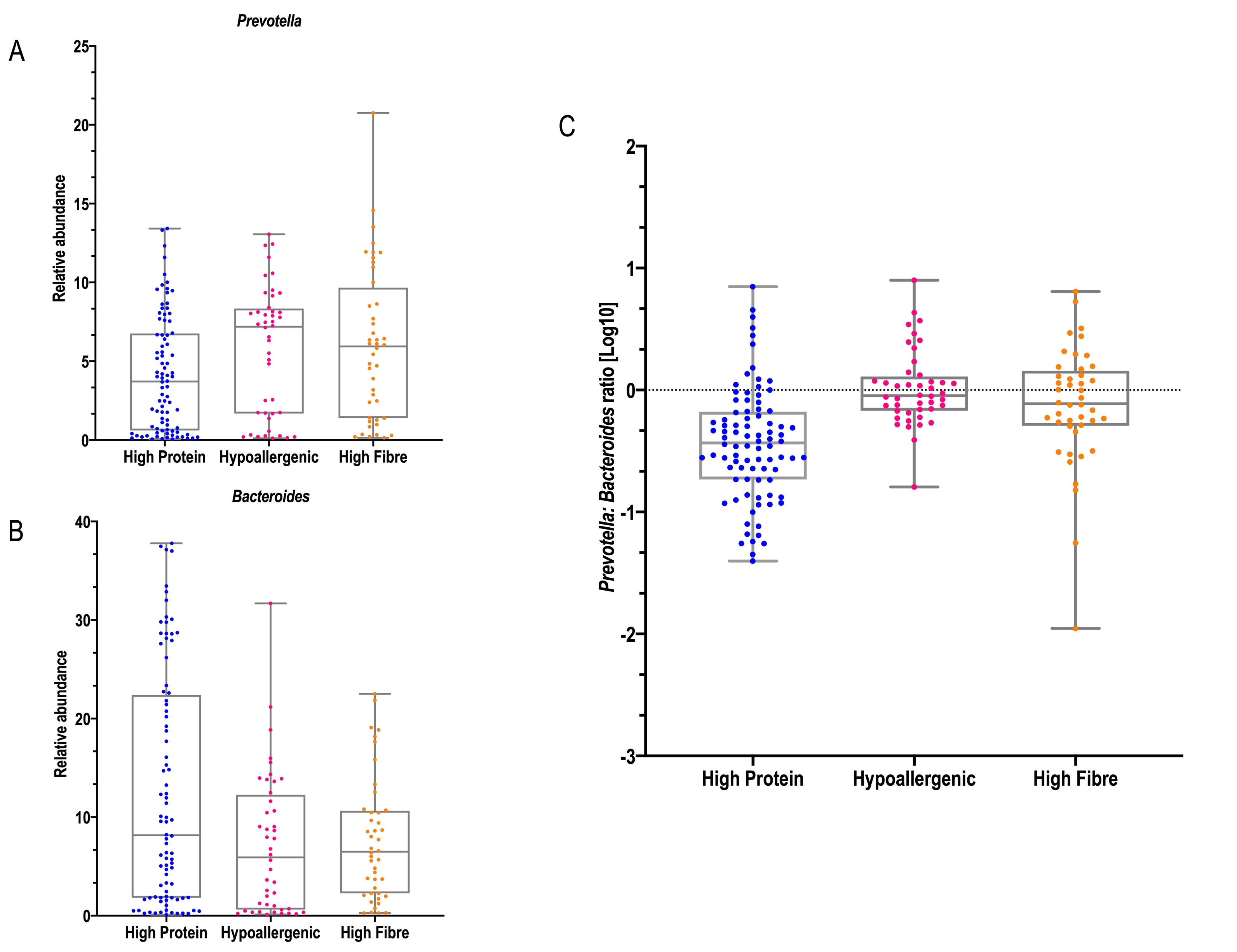
At lower phylogenetic levels, an overrepresentation of members of the families Clostridiaceae and Peptostreptococcaceae were also found in the samples from dogs fed with the diet C. Clostridium is important for lysine and proline utilisation by the host via fermentation in the colon, while Peptostreptococci drive tryptophan and glutamate catabolism [52]. In people, an exclusively meat-based diet is frequently associated with high levels of bile-tolerant bacterial species like Bacteroides and low levels of Prevotella [43]. Of interest, Clostridiaceae and Peptostreptococcaceae were only enriched in the washout period, whereas the Bacteroidaceae family was enriched during baseline, emphasising the effects of previous diet in the microbiota profile.
Studies that have evaluated the impact of low-fibre/high-protein meat-based raw diets in the gut microbiome of healthy dogs [23, 24, 53] have shown an overall decrease in the abundance of Firmicutes, including genera Peptostreptococcus and Faecalibacterium; and in Bacteroides and Prevotella (phylum Bacteroidetes). Conversely, other bacterial taxa were found to increase in abundance, including Proteobacteria and Fusobacteria (genus Fusobacterium) [23, 24], and two genera from phylum Firmicutes (Lactobacillus and Clostridium) [23, 53].
Although previous studies have identified increased levels of Enterobacteriaceae in dogs fed raw diets, we did not see enrichment of this bacterial group during this dietary intervention [53].
Another difference of the high protein diet compared to the other diets was the percentage of fat. Studies have reported that an animal-based diet high in fat (independent of protein) resulted in substantial changes to the microbiota and metabolites produced [4, 43, 54]. Increases of bile-tolerant organisms [4] and members of the Proteobacteria family [55] and in the Firmicutes: Bacteroidetes ratio [56]. However, fat is not a homogenous macronutrient, and the structure and function can vary significantly among the type of fat [2]. Likewise, in an all meat diet fed in the manner in our study individual dogs may receive differing quantities of fat due to different distribution within tissue. Further studies need to be done to unravel the precise effect of fat on the gut microbiome in dogs.
Fibre has historically been classified as either soluble or insoluble, but plant cell walls often contain both, and this distinction does not always predict physiological function [2, 57]. Although an agreement has not been reached, and several classification systems have been proposed, the US Codex Alimentarius Commission introduced the following definition of dietary fibre in 2009: ‘carbohydrate (CHO) polymers with ten or more monomeric units, which are not hydrolysed by the endogenous enzymes in the small intestine of humans and belong to the following categories: (a) edible CHO polymers, naturally occurring in the food as consumed; (b) CHO polymers, obtained from food raw material by physical, enzymatic, or chemical means; and (c) synthetic CHO polymers [58].
The high-insoluble fibre diet (Diet B) used in this study contains 25.5% (dry matter [DM]) insoluble fibre and 1.9% DM soluble fibre; the total dietary fibre is therefore 27.6 % DM and crude fibre 16.4% DM. This type of diet is used for conditions such as weight loss, diabetes mellitus, chronic pancreatitis and historically for conditions liked colitis. Most standard canine diets fed for maintenance in adults contain crude fibre around 1.5-5 % DM. Diet B in our study induced an enrichment in bacteria belonging to the Firmicutes and Actinobacteria phyla. However, at family level, Prevotellaceae (belonging to the Bacteroidetes phylum) was also enriched. This agrees with human studies, where it has been found that increased levels of Prevotella are associated with a plant-based diet rich in fibre, simple sugars, and plant-derived compounds, as they harbor genes for cellulose and xylan hydrolysis [43, 59].
Obligate anaerobic bacteria (phyla Firmicutes and Bacteroidetes) encode a variety of enzymes for hydrolysing complex carbohydrates not digestible by the host, such as plant cell wall polysaccharides and resistant starch, which constitute most dietary fibres [60, 61]. These components enter the large intestine and undergo microbial breakdown and subsequent fermentation. The major end products of microbial fermentation are short chain fatty acids (SCFAs), including butyrate, propionate, and acetate [62]. They are rapidly absorbed by the intestinal epithelial cells where they are involved in several cellular and regulatory processes with only 5% excreted in faeces [63, 64]. Butyrate, mainly produced by Firmicutes, constitutes the main energy source for the epithelial cells [65] and plays an important role in brain function [66]. It is also known for its anti-cancer [67] and anti-inflammatory properties [63] and for its role in the development of the intestinal barrier [68].
Hypoallergenic and/or hydrolysed diets are used frequently in dogs for the treatment of putative food allergies and chronic enteropathy [33, 36, 69]. The main difference between a commercial dry diet designed for healthy dogs and a hypoallergenic diet is that the latter is composed of hydrolysed protein that decreases the probability of an immune response to protein dietary components [33]. The diet used (Diet A) is based on hydrolysed poultry, and although has lower fibre content and higher fat content than Diet B, it is similar in overall macronutrient composition to commercial maintenance diets. Evaluation of the effect of the hydrolysed diet did not show overrepresentation of any member at the phylum and family phylogenetic levels, in comparison with the other two diets. Potentially, dietary impact of hypoallergenic diets on the gut microbiota could be at functional level and not necessarily at taxonomic levels. It could lead to changes in bacterial metabolites that can promote the production of immunoregulatory metabolites, which interact with the host immune cells to promote non-responsiveness to innocuous luminal antigens (SCFAs) [62], stimulate secretory immunoglobulin A (sIgA) and β-defensins production; modulate the cytokine response or lead to an improvement of the intestinal barrier, ameliorating the clinical signs in dogs with intestinal inflammation [70]. Further studies, assessing function and strains could help us to elucidate the relevance and the role of these microbiota changes in the gut. It is interesting that these changes were different and independent from the high fibre diet, which suggests a different mechanism of action. Although we used PICRUSt to predict community functional’s capabilities, we could not find an effect based on the type of diet or diet sequence.
Recent studies evaluating the effect of a hypoallergenic diet on the gut microbiome in healthy dogs and in dogs with food-responsive enteropathy showed that the impact of the diet was minimal in the microbial composition as well as in the metabolome [16, 37]. In these studies, dogs were fed with commercial maintenance diets before the introduction of the new diet, whereas in our study the baseline diet was meat-based, which could potentially have an influence on the results. Also, the percentage of fat differed among hypoallergenic diets, with our diet being slighter higher in fat percentage.
We also saw that the magnitude and nature of the changes induced by the high fibre and hydrolysed diets varied according to the diet sequence. The initial bacterial composition, the fact that bacteria form a metabolic network and cross-feed each other and that there is significant heterogeneity within bacterial species in their ability to digest different types of fibre [2, 71] add complexity to the diet-microbiota interaction. In people, particularly in the case of fibre, it has been shown that an individual’s baseline microbiota harbors predictive potential with regards to the effect of dietary constituents on the host [72]. Also, we should take into consideration that the proportion of one macronutrient to the total energy intake inherently influence the contribution from other macronutrients. Thus, the effect of a change in one macronutrient on the faecal microbiota is therefore a result of the combinatory effect of all the macronutrients [73]. Both, the decrease or the increase of a nutrient can contribute to the changes seen in a diet.
In our study, we observed that the ratio of Prevotella to Bacteroides was higher with the hydrolysed and high-insoluble fibre diets compared to the high-protein diet.. In accordance with this, it has been reported that a high fibre diet correlates with a microbiome consisting of polysaccharide-utilizing microbiota with lower protein fermentation products and fewer Bacteroides and Clostridia [74, 75]. However, when we analysed the families using the Dirichlet model, we observed that Prevotellaceae was only higher in the high-insoluble fibre diet and only in the ACB sequence, whereas members of the Bacteroidaceae family were higher in the high-protein diet but only during the baseline period.
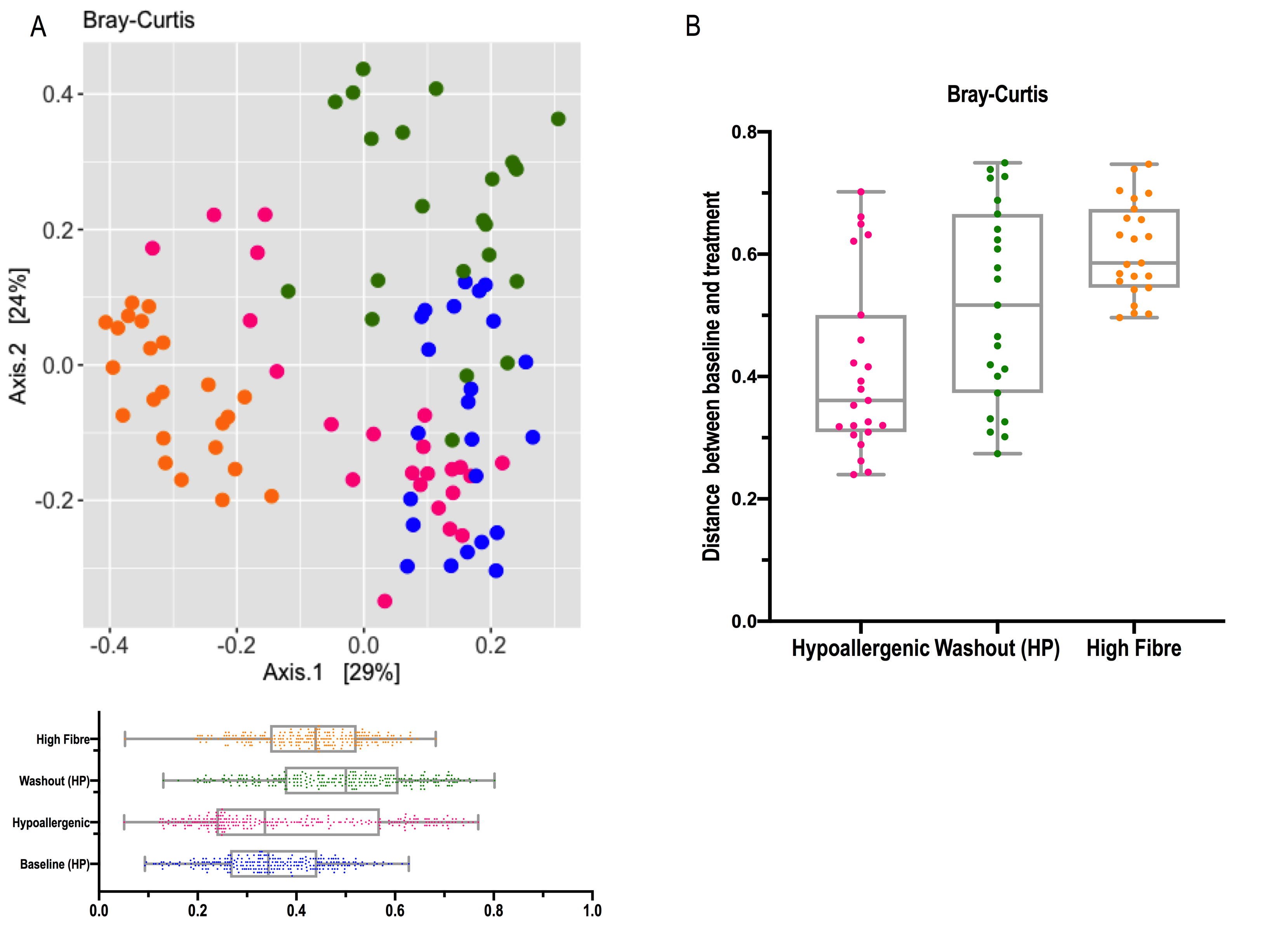
Finally, assessment of the gut microbiota during the washout period showed that the gut microbiota of dogs did not revert to their original phylogenetic structure after six weeks. Previous studies in dogs have reported adaptation periods varying from 10 days to 4 weeks [5, 7, 8, 24, 76]. In our study, although the washout period was longer than previously reported, changes in the composition of the gut microbiota persisted over time. This was evidenced by sequence and diet effects and by differential results in bacterial abundance between baseline and washout periods. These changes could be permanent or there is a possibility that more time is required with the original diet to return to baseline levels. The intestinal microbiota is resistant to most environmental influences, returning rapidly to its pre-treatment state, particularly for short-term interventions [4]. Furthermore, studies have shown that it seems that long term improvements to dietary habits may be required to achieve permanent changes in the gut community structure [43]. However, this can depend on the magnitude and duration of the change [4, 77]. The credible interval for the phyla and for the 20 most abundant families in each diet was quite broad. This could be due to small number size, high between-sample variability, substantial uncertainty in the taxa-specific effects, among other factors. For most families the predicted abundance was relatively low, making effects on those taxa difficult to detect.
The limitations of this study were the presence of only one breed, age (although they were evenly distributed in both groups), and potentially the manufacturing process of the commercial diets themselves could also have influenced the gut microbiota. Additionally, the alternate day feeding pattern of Day C could have influenced microbiome composition independently of the protein/digestibility [78,79]. Furthermore, day to day variations in microbiota occurs and in our study, faeces were only collected at a set time point [41]. Pooling samples over a collection period of several days may have been more beneficial to average out day-to-day variability but would have added more complexity to the analysis.
In addition, evaluation of microbial composition together with functional analysis (metabolomics, transcriptomics) would offer a better insight in the total effect of diet [71]. Different microbiomes have different potentials for producing certain metabolites, depending on the metabolic capabilities and metabolic interactions within the population. The fact that a bacterium harbours a gene does not imply that the gene is expressed. In the presence of different energy sources, bacteria may express genes to produce one, a group or several of these enzymes, depending on the environmental context [71]. Future studies could combine several approaches to elucidate the influence of the diet-microbiota interaction on host biology.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}