Cardiid phylogeny
This is the first phylogenetic study using transcriptomic data that aims to resolve the relationships between the two cardiid subfamilies which contain photosymbiotic taxa. The best corroborated hypothesis supports a sister relationship between Tridacninae and Fraginae, both recovered as monophyletic (Fig. 3). Other than the position of Tridacninae, the placement of all other cardiid subfamilies is consistent with the most recent multi-gene Cardiidae phylogeny comprising 110 species [19].
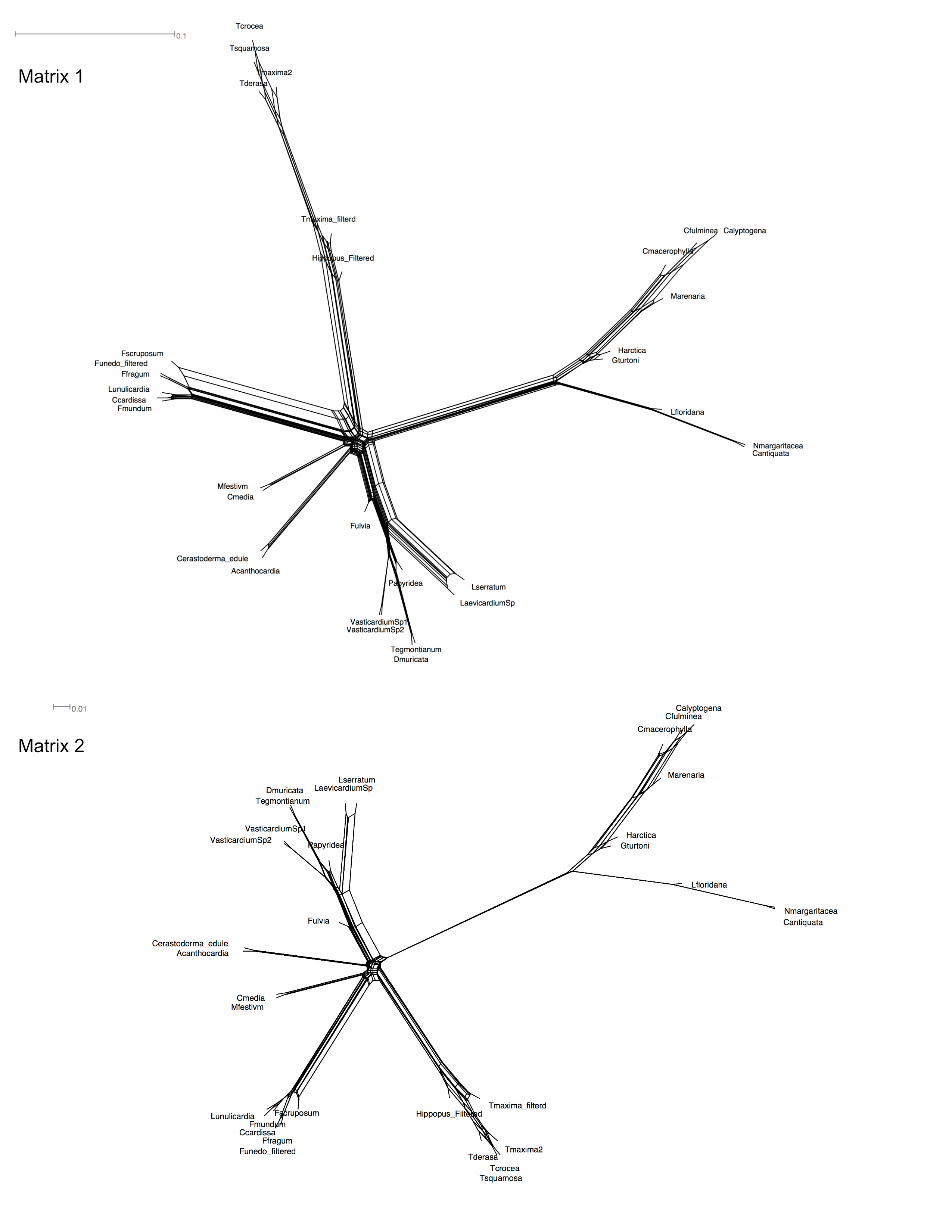
Our analyses highlighted the pervasive challenge to reconstruct a well-resolved and highly supported cardiid phylogeny. The fundamental reconstruction difficulty stems from the diversification process of Tridacninae, photosymbiotic, and non-symbiotic Fraginae. Our results show that the branches leading to the three crown groups are long and subtended by very short internodes, indicating the divergence among these groups was rapid. These ancient but fast diversification events are inherently difficult to resolve [73]. Some of the major problems include: not enough data to resolve the nodes; molecular data not variable enough at the appropriate level; strong conflicts among genes; and inadequate substitution models [73]. We discuss these concerns in the following paragraphs.
Firstly, current studies on cardiid phylogeny may suffer from some level of data limitation. For example, although Herrera et al. 2015 [19] had excellent taxon sampling through Cardiidae, the gene coverage was low. In the present study, taxon sampling was limited due to the need to obtain high quality transcriptomes, which requires freshly collected specimens. These constraints could be overcome by incorporating DNA-based phylogenomic approaches (e.g., RADseq, exome capture, etc.) using well-preserved museum specimens.
In addition, some of the data may lack the appropriate level of resolution. For example, the ML Analysis on Matrix 2 resulted in poorly resolved topologies with low bootstrap support values. This observation where large, albeit more incomplete matrices provided better resolution than small and more complete matrices is not unique to our dataset (e.g., [24, 45, 74]). It is likely influenced by gene choice in the more complete matrix [74]. If a gene is present in more taxa, it is likely to be relatively conserved; hence lacking enough genetic variation to resolve rapid divergence, as seen in some of our individual gene trees. These very conserved/slow-evolving genes might be overrepresented in a smaller matrix, but contribute minimally to any phylogenetic resolution.
Gene conflicts are also apparent in our dataset based on the supernetwork and submatrices analyses, in part may be explained by incomplete lineage sorting at these rapid-divergence nodes. In addition, genes with different rates of evolution gave rise to divergent topologies. Some faster evolving genes (e.g., Matricis Q-V) produced clearly problematic topologies, such as placing photosymbiotic Fraginae as sister clade to all other cardiids. These rapidly evolving genes might be subject to more genetic saturation and could be under the influence of strong selection, all of which hinder their ability to resolve deep phylogenetic nodes and conflict with other more informative genes.
Lastly, sequence compositional heterogeneity is known to be especially problematic for inferring short internal nodes [75]. However, based on the low level of composition heterogeneity in our dataset and the Dayoff recoding analysis, it is unlikely to produce significant bias in the results.
In sum, we have found that overly conservative genes and fast evolving genes don’t provide informative resolution to the nodes of interest, a phenomenon observed in other taxonomic groups [76]. These genes generate conflicting topologies and could be responsible for the observed low bootstrap values. On the contrary, ML analyses on the large matrix, moderately evolved genes, as well as the Bayesian analysis, all support a sister relationship between Tridacninae and Fraginae.
Implications on photosymbiosis evolution
Our analyses support two independent origins of photosymbiosis in Cardiidae. This result should be robust to phylogenetic uncertainties, because a non-sister relationship between Tridacninae and Fraginae will only reinforce the independent evolution scenario. Our ancestral state reconstruction was model-based; an alternative parsimony-based analyses would deem “two independent origins” or “one origin and one loss (in the non-symbiotic Fraginae clade)” equally likely. However, the latter scenario is not well supported because fossil record indicates that visible shell adaptations in both Tridacninae and Fraginae appeared after Late Oligocene [3, 77]. If the common ancestor of the two subfamilies is photosymbiotic, it would imply that photosymbiosis was established in late Cretaceous but persisted more than 30 Ma without any apparent shell adaptations. It is much more probable that there are two separate origins of photosymbiosis with shell adaptations evolved shortly after.
The repeated evolution of bivalve photosymbiosis suggests its adaptive advantage. The relatively rapid crown group diversification, coupled with morphological responses [3], is consistent with criteria for adaptive radiation - the generation of new species exhibiting pronounced morphological divergence over relatively short timeframes, typically in response to new environmental conditions [78]. In photosymbiotic cardiids, the species number is modest compared to other documented examples. However, this might be due to the lack of systematics research in these groups, as more studies have started to reveal their hidden diversities (e.g., [79]).
As for most cardiid lineages, Tridacninae and Fraginae have inferred ancestral distributions that span the Indo-Pacific; Tridacninae has a wider ancestral range, reaching the western temperate northern Pacific [19]. Given that the origin of photosymbiosis in both subfamilies overlaps with the expansion of modern reefs, it is likely that the formation of shallow marine habitat in the Indo-Pacific is a key environmental driver for bivalve photosymbiotic adaptation. This is further supported by the fact that photosymbiotic fraginaes has a sister non-symbiotic lineages that still inhabits deep sandy bottoms [7], possibly representing the ancestral ecology of these bivalves before they shifted to shallower habitat. Kiessling 2009 [72] proposed a fundamental question regarding the formation of reef biodiversity - Have reef taxa originated within reefs or have they evolved somewhere else then moved into reef habitats? Our results indicate that at least for photosymbiotic bivalves, they have likely originated and diversified within the reef habitats. More biogeographical, palaeontological and phylogenetic data are certainly needed to further corroborate this point.
Both Tridacninae and photosymbiotic Fraginae are thought to exhibit phenotypic adaptations to benefit the symbionts, making some species’ shell and mantle morphologies drastically different from typical cardiids (Fig. 1). What is striking, and little mentioned until now, is the divergent morphological trends in photosymbiotic fragines contrasted with the uniform morphological trends in Tridacninae, essentially two different responses to expose symbionts to optimal irradiance in sister lineages. Previous studies of adaptive radiations have well highlighted examples of divergent morphological responses to newly available niches (e.g., [80]), as seen in fragines. But uniform morphological responses, as seen in tridacnines, are less documented. Both strategies have advantages. While divergent morphologies provide highly specialized adaptations to different fine-scale niches, an uniform/static morphology may enable the lineages to become broadly adapted generalists to mediate environmental fluctuations [81]. The different morphological evolution patterns in photosymbiotic cardiid suggests that in the acquisition of a key innovation, historical morphological contingencies (e.g., opaque heavy shell, unexposed tissues, infaunal habit) and common ancestry do not predict the directionality of morphological evolution, not even for sister lineages that use the same ecological strategy to adapt to similar habitats.
Despite the versatile shell and mantle morphologies in Fraginae and Tridacninae, their symbiont-containing tubular system share striking similarities. Both stem from the digestive system of the hosts and form tertiary tubular networks [13, 16]. The only other similar molluscan structures are found in some marine gastropods who temporarily maintain algae or chloroplasts in their tissues [82]. The development of giant clam tubules are only triggered by the presence of symbionts [14], indicating the acquisition of photosymbiosis is a highly interactive process between hosts and symbionts.
It has long been hypothesized that different photosymbiotic bivalves express homologous genes to build the tubular system, in response to similar symbiont signals [16]. The newly-found sister relationship between Tridacninae and Fraginae lend further support to this theory, suggesting that genes homologous to the tubular-formation ones are ancestral to the two lineages. It is even possible that the genes are ancestral to bivalve and gastropods, as the latter can form similar anatomical structures. Given that the different mollusk lineages evolved photosymbiosis independently, it is likely that the gene/anatomical level similarities are generated from parallel evolution. That is, each lineage independently coopted similar genetic mechanisms for generating symbiont-containing structures. Molecular level parallel evolution has been shown in photosymbiotic systems. For example, both corals and giant clams repurpose the expression of the vacuolar H+-ATPase gene (VHA) to facilitate their carbon concentrating process and promote algal photosynthesis, even though the two lineages are very distantly related [59, 60]. Our analyses further support that VHA is a conserved gene, ancestral to bivalves, corals, and other animals. Photosymbiotic bivalves don’t possess any “special” version of VHA; the amino acid sequence is the same as the ones found in non-symbiotic taxa. It is more likely that the adaptation to photosymbiosis is realized at the regulatory/expression level, where VHA is highly expressed in tissues that harbor symbionts. In contrast with the relatively labile shell and mantle adaptations, it is possible that the evolution of host metabolomic and developmental adaptations are more constrained genetically, resulting in similar mechanisms in diverse animal groups. More in-depth studies on the molecular mechanisms behind photosymbiosis are needed to gain better insights.
To further investigate macroevolution of bivalve photosymbiosis, a better understanding of potential photosymbiotic fossil taxa is essential. However, they are challenging to identify in the fossil record based on shell morphology alone [2]. Although photosymbiotic bivalves possess a suite of morphological traits to enhance light exposure, almost all characters used to support photosymbiotic condition are found in modern non-photosymbiotic bivalves. Examples include permanent shell gaping (Family Limidae, Galeommatidae), enlarged mantle (Galeommatidae), or transparent shells (Placunidae, Pinnidae). The only photosymbiotic-exclusive shell character is the shell “window” microstructure [17], but many photosymbiotic bivalve species do not possess it, and similar features have not been found in fossil taxa. Similarly, other identifiable photosymbiotic ecological traits (shallow water distribution, fast growth, reclining habits) are not unique to photosymbiotic bivalves either [2]. Therefore, alternative methods need to be developed (e.g. better isotopic metrics, metabolite signatures) if we wish to fully understand the evolutionary dynamics of bivalve photosymbiosis.
Lastly, further characterization of host-symbiont diversity and interactions are essential for developing a full picture of animal-algal photosymbiosis. A comprehensive documentation of host-symbiont biodiversity can provide insights into photosymbiotic mechanisms in diverse habitats and with different organismal complexities. For example, Li et al. 2018 [8] demonstrated that fragine species at varying water depths have different dependencies on algal photosynthesis, with deeper host species utilizing less photosynthetically derived carbon and exhibiting less shell modifications. Therefore, it is highly likely that host-symbiont interactions and reliance vary greatly among host-symbiont pairs, depending on symbiont types, as well as degrees of host adaptations. Li et al. 2018 [8] summarized known symbiont diversity in giant clams and showed that several fragine species harbor symbionts from the genus Cladocopium, which also occupies cnidarians, giant clams, and foraminifera. However, this by no means captured the full spectrum of potential symbiont diversity in photosymbiotic bivalves, especially when new host species are discovered regularly (e.g., [79]). In most photosymbiotic bivalve systems, the roles symbionts play in host ontogeny, reproduction, response to environmental fluctuation, etc. are unexplored, all of which require our continued effort to identify and characterizing existing photosymbiotic diversity.
{kind=link}
{kind=link}
{kind=link}