Human THAP9 forms a homodimer.
DmTNP has been reported to oligomerise as a prerequisite for active P element transposition (15). However, it is not known if hTHAP9, the less studied homolog of DmTNP, forms oligomers. Thus, the possibility of hTHAP9 forming oligomers was probed by independently tagging the protein with FLAG and HA tags (THAP9-F, THAP9-H) and observing if these differently tagged proteins colocalise and form a homodimer.
Immunostaining and immunoblot analysis established that THAP9-H and THAP9-F expressed optimally, 24 and 48 hours post transfection (into HEK293T cells) respectively (Fig. 1A). Interestingly, they were found to colocalise in the nuclei, both 24h (Fig. 1B) and 48h post co-transfection (Fig. 1C). The colocalisation of THAP9-H and THAP9-F was statistically significant (P value < 0.0001, Fig. 1E) when compared to control (lipofectamine treated cells, Fig.1D)
Since THAP9-H and THAP9- F showed optimal expression at different time points, pull-down assays were performed instead of co-transfecting both constructs followed by co-immunoprecipitation. When THAP9-F was used as a bait, it could pull down THAP9-H (Fig. 1F, lane 1). THAP9-H bound to beads was run as a positive control (Fig. 1F, lane 2).
Thus, colocalisation and pull-down experiments establish that hTHAP9 appears to undergo homo-oligomerisation in the nucleus.
Prediction of amino terminal leucine rich coiled coil regions in DmTNP and hTHAP9.
We have previously identified an evolutionarily conserved, leucine rich, alpha helical coiled coil region, downstream of the amino terminal THAP domain of most human THAP proteins including THAP9 (27). Since coiled coil regions mediate protein oligomerization (33), it was predicted that the identified coiled coil in human THAP9 may be involved in oligomerisation.
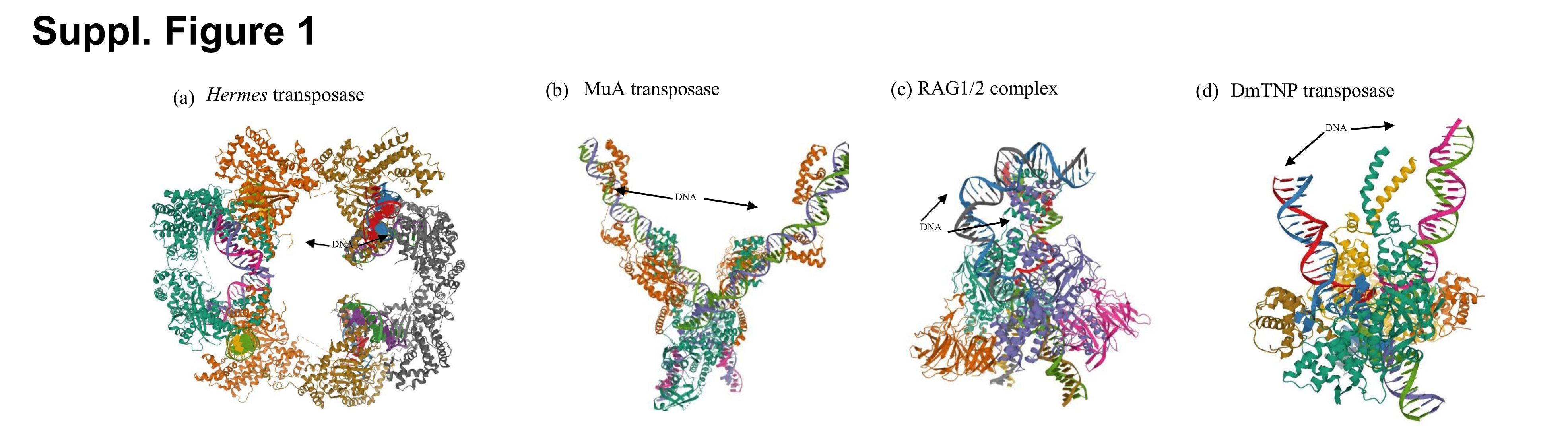
DmTNP, which is homologous to hTHAP9, is known to exist in different oligomeric forms at different steps of transposition; as a tetramer bound to DNA in the PEC/CDC (15) and as a dimer in the post-transposition STC (17). However, the recent cryoEM structure of the dimeric DmTNP STC could not resolve the flexible amino terminal DNA binding domain and adjacent dimerization domain (residues 1-150) (17) and thus the role of these domains in DmTNP oligomerisation is not clear. Hence, we decided to probe if there was an additional oligomerisation region in the amino terminal region of DmTNP.
COILS analysis predicted that multiple short regions throughout the length of DmTNP had a higher probability (>0.5) of forming coiled coils (Fig. 2A, top panel). In hTHAP9, some short regions in the centre were predicted to have higher probability (>0.5) while the amino terminal region had modest probability (>=0.3) of forming coiled coils (Fig. 2A, top panel). JPRED predicted that these putative coiled coil regions had a very high tendency of forming alpha helices till residue 140 for DmTNP and till residue 176 for hTHAP9 (Fig. 2B). Interestingly, a strong alpha helical region was independently predicted till residue 165 in hTHAP9.
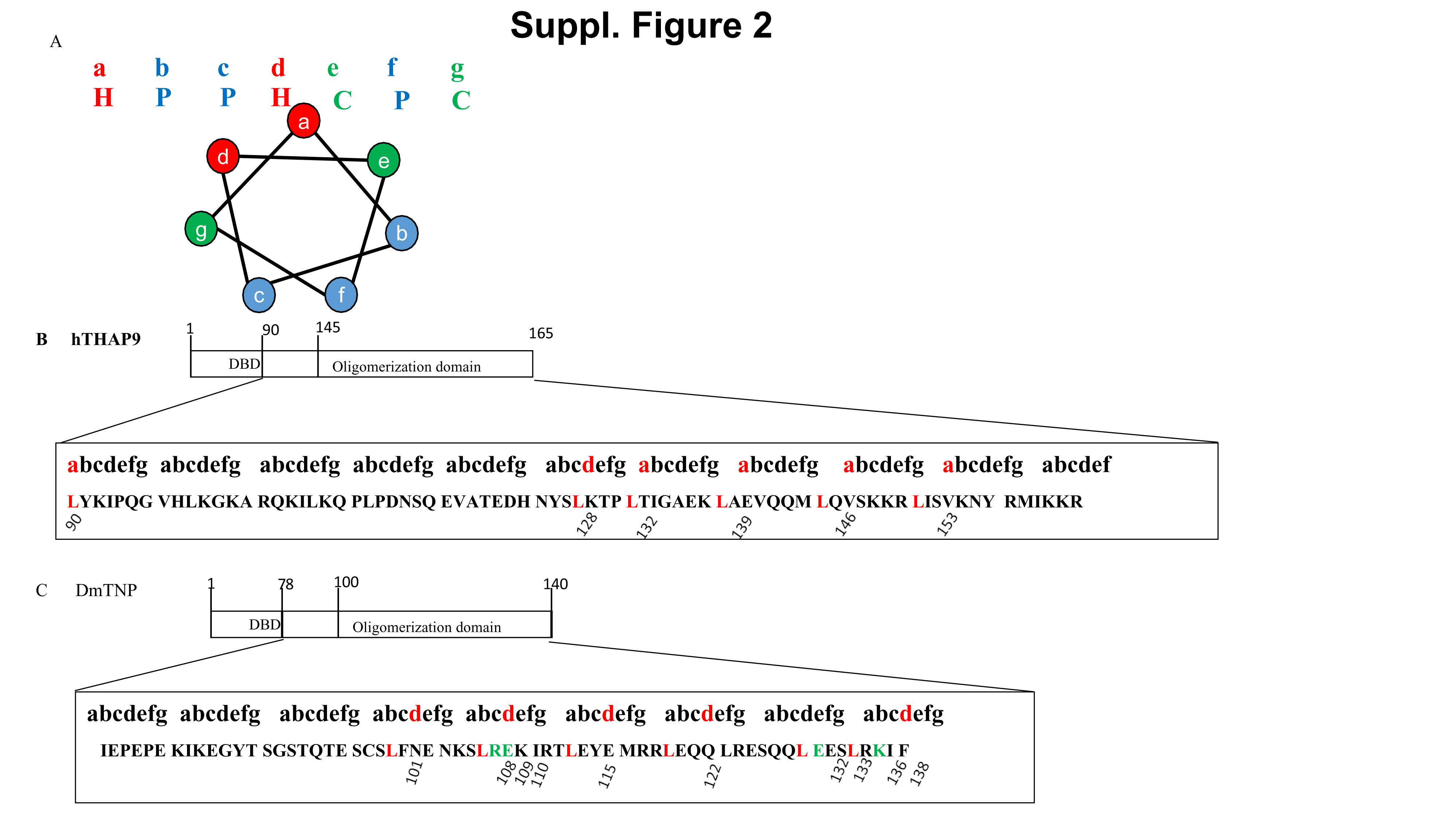
Coiled coils are characterised by “heptad repeats” (repeated pattern of seven amino acids, a-g), in which the a and d positions are non-polar/hydrophobic residues (like Leu) while the e and f positions are charged amino acids (Suppl. Fig. 2A). During protein oligomerisation, the side chains of hydrophobic residues on one monomer mediate ‘knob into holes’ packing with similar regions from another monomer. This leads to the formation of homo-oligomers (33), characterized by amphipathic alpha helices of individual monomers twisting around each other and enclosing a hydrophobic core. The regions immediately after DBD till the end of the predicted coiled coil region (hTHAP9: 90-165 residues; DmTNP: 78-140) were aligned to the heptad repeat pattern. Interestingly, some leucines in both proteins aligned to the a and d positions (Suppl. Fig. 2B, 2C) (27), consistent with previous reports (19, 27).
To investigate if the predicted alpha helical regions (from Fig. 2A; DmTNP: residues 100-140 and hTHAP9: residues 145-182) could form amphipathic helical structures via electrostatic interactions amongst themselves, DRAW COIL analysis was performed. Multiple electrostatic repulsive interactions were predicted in both these regions as seen in Fig. 2C.
Nuclear expression of truncated DmTNP and hTHAP9.
Based on our bioinformatic analyses (COILS, JPRED, DRAW COIL; Fig. 2), we decided to experimentally investigate the role of the amino terminal DBD and leucine rich coiled coil regions in DmTNP (D =residues 1-140) and hTHAP9 (H1=residues 1-176 and H2=1-165). We constructed both HA- and FLAG-tagged carboxy-terminal truncated clones which included the amino terminal DBD (DmTNP- 1-77 residues, hTHAP9- 1-94 residues), the predicted nuclear localization signal (NLS) (DmTNP- 60-69 residues, hTHAP9- 150-168 residues) and the predicted coiled coil regions [DmTNP- 100-140 residues, hTHAP9- 145-165 (H2) or 145-176 (H1) residues] of each protein.
Each FLAG-tagged (F-H1, F-H2, F-D) and HA-tagged (H1-H, H2-H, D-H) truncated protein was individually found to express and localize to the nucleus of HEK293T cells (Fig. 3A and 3B). HEK293T cells treated with lipofectamine alone (bottom panels) confirm that there was no background signal. Western analysis demonstrated that all these constructs [approximate MW: 20 kDa (H1), 19 kDa (H2), 17 kDa (D)] expressed optimally 48h post-transfection (Fig. 3C and 3D).
Nuclear co-localisation of truncated DmTNP and hTHAP9 suggests homo-oligomerisation
Truncated differentially tagged DmTNP (F-D, D-H) or hTHAP9 (F-H2, H2-H or F-H1, H1-H) were found to co-localise in the nucleus. Briefly, immunostaining of HEK293T cells co-transfected with either F-H1 and H1-H (Fig. 4A), F- H2 and H2- H (Fig. 4B) or F-D and D-H (Fig. 4C) demonstrated that truncated DmTNP or hTHAP9 appear to homo-oligomerise in the nucleus. The colocalisation was statistically significant with P value < 0.0001 (Fig. 4E).
The observed nuclear co-localisation may be due to the spatial overlap or proximity of these truncated proteins in the nucleus. To further probe this, we performed proximity ligation assays (PLA), which can detect possible interactions between two proteins, less than 40 nm apart, as foci (red). Briefly, HEK293T cells co-transfected individually with (Fig. 5A: F-H1 and H1-H; F-H2 and H2-H or F-D and D-H) were fixed and used for PLA using two different antibody pairs. Counting PLA foci (red) established that F-H1 and H1-H; F-H2 and H2-H as well as F-D and D-H occur in close proximity (P <0.0001) when compared to control (lipofectamine treated HEK293T cells).
Although, some PLA foci were detected in the extranuclear region, (Fig. 5A)
H1, H2 and D homo-oligomerisation was more prevalent in the nuclei (P value <0.0001) than the cytoplasm. The preferred nuclear interaction could be due to the presence of the DBD and predicted NLS in the truncated constructs.
Homo-oligomerisation of truncated DmTNP and hTHAP9 confirmed by co-immunoprecipitation
To further confirm if the observed co-localisation (Fig. 4 and 5) was due to the interaction of the truncated proteins, we performed reciprocal co-immunoprecipitation assays on whole cell lysates obtained from HEK293T cells co- transfected with F-H1 and H1-H, F-H2 and H2-H or F-D and D-H. Immunoprecipitation of F-H2 and F-D using HA antibody and the reciprocal immunoprecipitation of H2-H and D-H using FLAG antibody, confirmed the interaction between F-H2 and H2-H (Fig. 6A) and F-D and D-H (Fig. 6B).
Interestingly, the reciprocal co-immunoprecipitation of F-H1 and H1-H was very weak (Suppl. Fig. 3). H1 (residues 1-176) has 11 additional amino acids (residues 166-176) when compared to H2 (residues 1-165). Thus, this suggests that the 1st 165 residues of hTHAP9 (H2) may constitute a minimal amino-terminal oligomerisation domain. It is also intriguing to note that the addition of residues 166-176 significantly abrogate the oligomerisation, without affecting protein expression.
Role of Leucines and oppositely charged amino acid pairs in oligomerisation
The predicted helical coiled coil regions of DmTNP and hTHAP9, which appear to be important for homo-oligomersation, are leucine rich (Fig. 2B, Suppl. Fig. 2).
DNA transposases such as IS911 have been reported to form multimers mediated by leucine rich coiled coil structures (7). The DmTNP inhibitor KP repressor has also been shown to dimerise via leucine rich coiled coil (19). Thus, we decided to investigate if the leucine rich regions (~40 residues long, Suppl. Fig.2) downstream of the amino terminal THAP domain in hTHAP9 (145-182 residues) (27) and DmTNP (100-140 residues) were involved in oligomerisation.
When the Leucine-rich regions of DmTNP and hTHAP9 were aligned to the heptad repeat pattern characteristic of coiled coils, several leucines in both proteins aligned to the a and d positions (Suppl. Fig. 2B, 2C). These residues (DmTNP: Leu at positions 101, 108, 115, 122, 132 and 136; hTHAP9: Leu at positions 90, 128, 132, 139, 146 and 153) were individually mutated to Ala (polarity and size similar to leucine) in the corresponding truncated proteins (D-H, H2-H respectively). The LtoA point mutants (on H2-H or D-H templates) were individually probed for disruption of interaction by reciprocal co-immunoprecipitation with F-H2 or F-D respectively. L90A_H2H and L101A_DH were not included in the co-IP experiments as they did not express well. Surprisingly, none of the LtoA point mutants of either hTHAP9 (Fig. 7A) or DmTNP (Fig. 7B) showed complete disruption of interaction. To check for the possible combined effect of all the Leucines, we created combined LtoA mutants for both hTHAP9 [H2(all)-H (L90A, L128A, L132A, L139A, L146A, L153A)] and DmTNP [D(all)-H (L101A, L108A, L115A, L122A, L132A, L136A)]. However, even the combined leucine mutants did not show complete disruption of interaction either for hTHAP9 (Fig. 7C) or for DmTNP (Fig. 7D).
It has previously been reported that double mutants of KP repressor protein (first 199 residues identical to DmTNP), in which L108 and L115 were mutated to either alanine, valine, aspartic acid and proline, retained the ability to dimerise in vitro (19). In a separate study it was found that mutating L101 and L122 in KP repressor protein, to either isoleucine and arginine or valine and histidine respectively, did not completely disrupt its ability to repress P element transposition in fruit flies (20).
NMR studies on THAP11 demonstrated that its carboxy terminal region (254-306 residues) formed a homodimer, which was partly stabilized by a salt bridge interaction between an oppositely charged pair (K299 and D300) that occurred at positions e and g of its heptad repeat pattern (34). Thus, we decided to investigate if the acidic-basic residue pairs (Suppl Fig. 2C, green residues) in DmTNP (R109 and E110, E133 and K138) played any role in its oligomerisation. Acidic residues E110 and E133 were mutated to basic residues i.e. Lys (E110K, E133K) while basic residues R109 and K138 were mutated to acidic residues i.e.Glu (R109E, K138E) in truncated DmTNP (D-H). However, it was observed that none of these point mutations (R109E, D110K, E133K and K138E) disrupted the interaction with F-D in reverse co-IP experiments (Fig. 7E).
Thus, leucine and charged pair mutations in truncated DmTNP and hTHAP9 do not disrupt homo-oligomerisation
Truncated DmTNP and hTHAP9 can oligomerise without the coiled coil region
The leucine residues in the coiled coil region did not appear to be important for hTHAP9 and DmTNP oligomerisation. Thus, we decided to delete the predicted oligomerisation region (DmTNP: 100-140 residues, hTHAP9: 145-165 residues) in both proteins to ascertain its importance. We constructed HA-tagged truncated clones which included the first 144 residues of hTHAP9 (H_Del-H) and the first 99 residues of DmTNP (D_Del-H;1-99 residues): both these constructs included the amino terminal DBD (DmTNP- 1-77 residues, hTHAP9- 1-89 residues) of each protein.
In a previous study, in vitro chemical cross-linking experiments using purified proteins reported that shorter DmTNP constructs containing the first 88 or 98 residues were unable to dimerise (18, 19). However, surprisingly, reciprocal co-immunoprecipitation demonstrated that neither H_Del-H (Fig. 8A) nor D_Del-H (Fig. 8B) lost the ability to homo-oligomerise. Thus, it appeared that both DmTNP and hTHAP9 were able to oligomerise in cells even without the predicted Leu-rich coiled region. More importantly, these surprising results suggested that the DBD and the region between the DBD and predicted coiled coil region may help mediate oligomerisation.
DNA mediates interaction between truncated DmTNP and hTHAP9
DNA transposases such as MuA and Hermes have been reported to form DNA-mediated homo-oligomers (8,9). Our surprising observations that both H_Del (Fig. 8A) and D_Del (Fig. 8B) were able to interact with F-H2 and D-H respectively, led us to hypothesize that these interactions could be mediated by DNA.
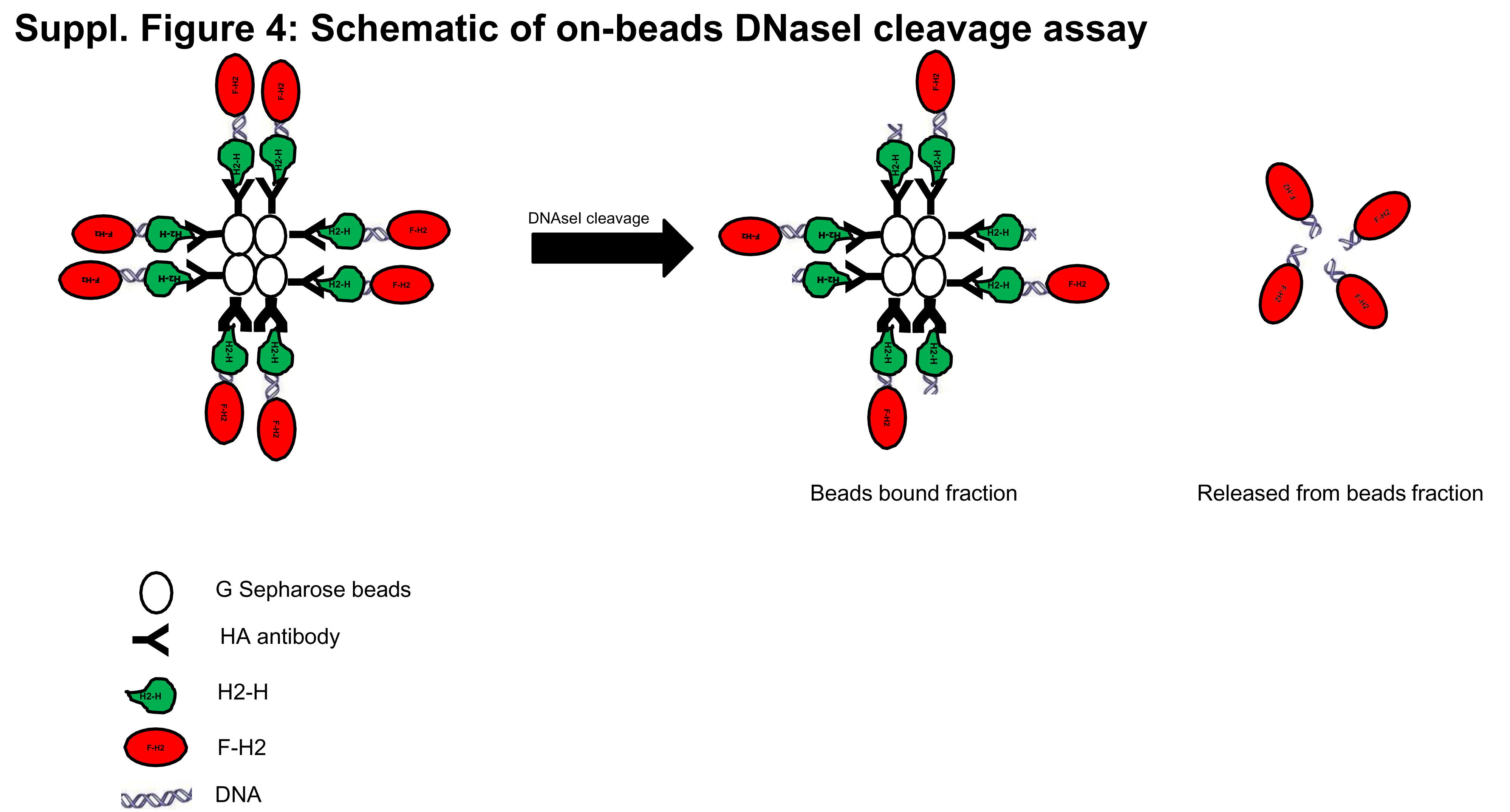
To investigate this further, on-beads DNaseI cleavage assays (Suppl. Fig. 4) were performed after reciprocal co-immunoprecipitation of F-H2 and H2-H (Fig. 8C) or F-D and D-H (Fig. 8D). If DNA mediates the interactions between each set of co-immunoprecipitated monomers bound to beads, the proteins should not remain bound to beads after DNaseI cleavage, as seen in Fig. 8C and 8D. On the other hand, if homo-oligomerisation is not DNA-mediated, DNase cleavage will not release the bound protein from beads. It is to be noted that DNase cleavage did not release all the bound protein (Lanes 2 and 3 in Fig. 8C and 8D); this may be due to incomplete DNase cleavage because of inaccessibility of the bound proteins to the enzyme or because the observed homo-oligomerisation of hTHAP9 and DmTNP truncated proteins is partially DNA mediated.
Human THAP9 may interact with Hcf-1 and other human THAP proteins.
Having established that hTHAP9 is able to homo-oligomerise, we decided to explore if the protein could also undergo hetero-oliogomerisation. Identifying hTHAP9’s protein interaction partners would also help understand its yet unknown cellular function. Interestingly, the STRING database (24) predicted that some human THAP family members namely THAP1, THAP10 and THAP11 may interact with hTHAP9. Moreover, hTHAP9 harbors a predicted Hcf-1 binding consensus motif [HBM, (D/E)HXY)] (25) between 123 -126 residues. Thus, we decided to investigate if hTHAP9 could interact with Hcf1, THAP1, THAP10 and THAP11.
THAP9-F was found to colocalise with H-Hcf-1 48h post transfection (Fig. 9A). Also, THAP9-H was found to individually co-localize with THAP1-F (Fig. 9B), F-THAP10 (Fig. 9C) and F- THAP11 (Fig. 9D) 24h post transfection. The co-localization of THAP9 with other protein interaction partners was found to be statistically significant (P value < 0.0001) when Pearsons’s R-value was compared to that of the control, as shown in Fig. 9E.
To define the minimal region required by hTHAP9 to interact with some of its partners, we checked if THAP1 or THAP10 could interact with truncated hTHAP9 (H2). Interestingly, H2-H did not co-immunoprecipitate with either THAP1-F or F-THAP10 (data not shown). This indicates that in addition to the amino terminal interaction regions described in this study, hTHAP9 may have additional carboxy terminal interaction domains which interact with THAP1 and THAP10.
Overall, we demonstrate that hTHAP9, like its homolog DmTNP, can undergo homo-oligomerisation, which is partially mediated by DNA. Deletion mutant, leu, H1Moreover, hTHAP9 can also hetero-oligomerise with several members of the human THAP family as well as Hcf1.
{kind=link}
{kind=link}
{kind=link}
{kind=link}