Morphology
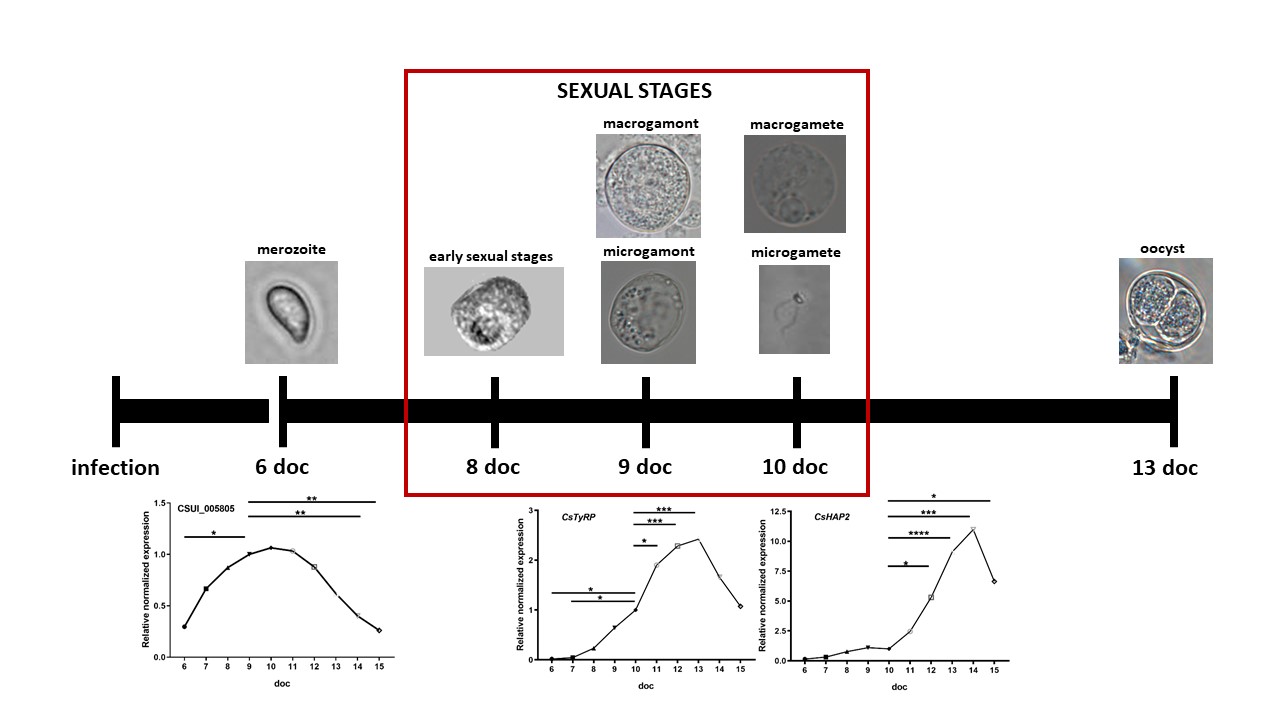
Sporozoite-infected cell cultures are a suitable model for producing all stages of C. suis in vitro. Free merozoites can be found in appreciable numbers from day 6 doc. Sexual stages occur in appreciable numbers between 8-11 doc and are mainly found outside the host cell. Early sexual stages were first detected at 8 doc. First gamonts could be located from 9 doc onwards, whereas first macrogametes and motile microgametes could be found a day later (Fig. 1). We estimated that the ratio of gamonts : early sexual stages was about 1 : 2. The first oocysts appeared 11 doc until 13 doc.
The sexual development of Coccidia, including C. suis, is not well characterized [8]. Although it is presumed that sexual stages are crucial in parasite development, little is known about their morphology, especially in vitro [6]. As the life cycle of C. suis takes longer in vitro (about 11 days) than in vivo (5 days) [4], it is possible to collect samples of every stage of C. suis development, hence making sexual stages available for further research.
Early sexual stages (immature gamonts) varied in form and size but their length was generally 15.6±0.5 µm (n=50) and their width 11.6±0.4 µm (n=50, Fig. 2a). Both micro- and macrogamonts were subspherical and had very similar diameter, however microgamonts were between 16.8±0.9µm (n=50), whereas macrogamonts were 16.6±1.1 (n=50) in diameter (Table 1). In light microscopy, unstained microgamonts were recognized by their large vacuole and motile microgametes inside (Fig. 2b, c). Both micro- and macrogamonts are often found in close proximity to each other (Fig. 2b, 2d, Additional file 1), and the egress of microgametes from microgamonts could be observed during a four-hour time frame. Each microgamont contained between 30-40 microgametes.
Scanning electron microscopy observations show that microgametes consisted of a small, spherical (3.5±0.5 µm, n=50) body with two opposing flagella of 11.2±0.5 µm length (n=50, Fig. 3a), which enabled the quick movement of the microgamete in search for a macrogamete. Macrogametes on the other hand were immobile, spherical with a smooth surface and had a diameter of 12.1±0.5 µm (n=50, Fig. 3b, Table 1).
Our study provides the first detailed in vitro characterization of sexual stages of C. suis. Previous in vivo studies already described immature micro- and macrogamonts in tissue sections of the small intestine [5, 36]. Early sexual stages are described as ovoid to elongate and smaller in size than those in vitro (immature microgamonts: 11.8 x 8.4 µm and immature macrogamonts: 9.4 x 6.5 µm) while the shape and size of mature micro- and macrogamonts are comparable to early sexual stages [6]. Other closely related coccidian species show similarities morphologies of sexual stages [37, 38]; however microgametes of T. gondii and Eimeria [17] have flagellae on the posterior end, whereas in C. suis they are clearly positioned on opposite sides, which might also affect their movement on the search for a macrogamete. With an in vitro system that allows for the collection and examination of mature sexual stages of C. suis, further studies on their properties and the fertilization process in Coccidia will be possible.
Genes linked to sexual stages
Identification of genes linked to the sexual stages is crucial to understand the developmental biology and the fertilization process of organisms with sexual development, including apicomplexan protozoa. Here, we analyzed eight of the genes with highest upregulated transcripts in microgametes, macrogametes and oocysts based on previous studies in Apicomplexa [11, 12, 27, 29, 31, 39–42]. To identify genes or their orthologues related to the sexual development in C. suis, we used the ToxoDB parasite database and determined four genes related to sexual stages and four genes related to meiosis and cellular division (see Table 2).
For microgametes, the orthologues of dynein light chain 1 (DLC1) and the male gamete fusion factor (HAP2), CsDLC1 (CSUI_000751) and CsHAP2 (CSUI_000472), clustered with the respective genes from closely related coccidian parasites. Sequence analyses revealed identities greater than 60% with T. gondii and Neospora caninum, signifying the close relationship of C. suis with these two species [42], and more than 36% with Eimeria necatrix (Table 2).
Two of the most highly transcribed genes found in macrogametes code for proteins involved in the formation of the oocyst wall. CsOWP1 (CSUI_006207) has more than 65% of identity with that of T. gondii (Table 2). Genome analysis of C. suis and T.gondii failed to identify orthologues for the GAM56 protein of Eimeria but revealed three low molecular weight hypothetical proteins possessing both a leader peptide and tyrosine-rich sequences. CSUI_001473 is an orthologue with 54% of homology with a gene coding for a tyrosine rich protein in Toxoplasma (TGME49_037080). Two of these three genes (TGME49_037080 and TGME49_087250) have peak expression levels in oocysts and the encoded proteins are incorporated as a part of the oocyst wall [40].
Among the genes with high expression in oocysts of Toxoplasma and Cryptosporidium [15, 40] we determined one coding for the meiotic recombination Rad51/Dmc1-like protein, and the orthologue in C. suis, CSUI_004539, had a 93% similarity to the gene of Toxoplasma (TGARI_272900) (Table 3).
Protein kinases have a role not only for signaling, but also during the transition states of cells [43]. Nima (“never in mitosis-gene A”)-related kinases of Plasmodium and Toxoplasma are involved in post-fertilization processes and in the meiosis [32]. We found three orthologues for Nima genes, one of them specific for male gametocytes, Nima1 (CSUI_004317) and two of them for female gametocytes, Nima2 and Nima4, (CSUI_003099 and CSUI_000744) and all three showed more than 40% of homology with the respective genes of Toxoplasma (Table 3).
The expression profiles of one gene related to asexual stages (merozoites) of C. suis (CSUI_005805 [44]), the four genes related to sexual stages and the four genes related to cellular division and meiosis were examined at different time points of cultivation in vitro in extracellular and intracellular parasites.
To evaluate the development of merozoites during cultivation and to test the suitability of the qPCR analysis for the detection of temporal changes in gene transcription, we included the uncharacterized merozoite-specific gene CSUI_005805. The level of the transcription for merozoites increased until it reached the maximum around 9–10 doc and then declined (Fig. 4). In previous studies, this gene showed a higher transcription in merozoites compared to sporozoites and in the present study, it showed higher levels during merozoite development compared to sexual development and oocyst formation. The transcriptional level increased throughout the entire merogony, indicating that this protein might be important for the establishment and/or growth of merozoites inside the host cell [44].
Transcript levels of genes related to sexual development in extracellular parasites were compared with 6 doc as a reference for merozoites released into the medium and 9–10 doc for extracellular gamonts/gametes. The transcription levels were eight to nine times higher in gamonts/gametes compared to merozoites (Fig. 5a), which agrees with the high level of upregulation demonstrated in RNA-seq analysis of sexual stages of Eimeria spp. [11].
Transcription levels of intracellular parasites were compared daily at 6–15 doc. As a reference point, we used day 10 because on that day first mature gametes were seen in vitro. The microgametes related genes transcripts reach peaks on 13–14 doc and the increase was 10– to 90–fold time higher compared to merozoites at 6 doc (Fig. 6) which is in agreement with results for RNA seq comparing merozoites from third-generation merozoites and gamonts of Eimeria [11]. The levels for DLC1 and HAP2 (Fig. 6a, c, d) transcripts on 13 and 14 doc were 30– and 60–fold higher, respectively, compared to day 6 with no detectable expression. Dynein proteins are part of the flagellum of microgametes and form part of the microtubule motor [45]. They are also involved in mitosis and meiosis, and are the major constituents of mitotic spindles, which are used to pull eukaryotic chromosomes apart [46]. As judged from the gene upregulation, these processes take place in C. suis in vitro between 9 and 15 doc. However, before that time point no transcription of DLC1 could be measured and the expression of DLC1 in Apicomplexa seems to be restricted to microgametes as described earlier [11].
In addition, we observed an upregulation of the gene expression of the fusogen HAP2 during 9–13 doc followed by a decrease (Fig. 6a, d). Its expression is also restricted to male gametes and its function in gamete fusion and during fertilization is extensively described for plants and unicellular eukaryotes [26, 47]. The results in C. suis correlate with previous reports on Eimeria and Toxoplasma. HAP2 is found in microgametes and unsporulated oocysts but not in sporulated oocysts or sporozoites [26, 27, 40, 48, 49]. Moreover, transcription of HAP2 was increased during the enteric development of T. gondii in the intestine of cats. HAP2 knockout parasites failed to fertilize and produce oocysts in vivo, and this supports the hypothesis that interfering with the fertilization process can be utilized in a transmission-blocking vaccine [29]. As for DLC1, expression seems to be restricted to microgametes.
Oocyst wall formation is a hallmark of coccidian development, and OWP and GAM-encoded proteins have previously been characterized in Eimeria and Toxoplasma as well as Cryptosporidium as constituents of the oocyst wall [29, 50, 51].
The GAM proteins EmGam56 and EmGam82 were identified as antigens that conferred protection against different species of chicken Eimeria due to their conserved nature. A subunit vaccine for immunization was previously developed from E. maxima gamont proteins and commercialized for the prevention of coccidiosis [9, 52].
No homologues for GAM genes were found in the C. suis genome or in the Toxoplasma database. However, oocysts of C. suis display the characteristic autofluorescence similar to other coccidia [17, 53] which is likely due to the dityrosine bonds formed between tyrosine-rich proteins present in the oocyst wall [11, 51]. A search in the proteome of Toxoplasma for predicted proteins with sequences rich in tyrosine identified five hypothetical proteins, and three of them presented highest levels of expression in the oocyst wall proteomic fraction [31] and were upregulated in oocyst transcriptomes compared to tachyzoites and bradyzoites [40]. We identified and analyzed an orthologue of one of them, TyRP. Gene transcripts of both proteins presumably involved in oocyst wall formation, OWP1 and TyRP, were upregulated until 13 doc and then steeply declined (Fig. 6b, e, f). While TyRP transcription could only be detected from 8 doc (Fig. Fig. 6b, f), low transcription levels for OWP1 were already found from 6 doc (Fig. 6b, e). indicating that merozoites present at this time point are probably already committed to a further development into macrogamonts, as described for E. tenella [17, 27].
In extracellular parasites, the RNA transcription of Rad51/Dmc1-like showed similar levels of transcription in parasites from 6 and 9–10 doc (Fig. 5b). In intracellular stages of C. suis, the RNA transcription levels of Rad51/Dmc1-like were rather constant from 6-12 doc except for an increase 13–14 doc (Fig. 7a, c) which probably corresponds to meiosis during the formation of the oocyst, since the RAD51/Dmc1-like gene codes for a protein of the Rad51 family which assists in repair of DNA double strand breaks during mitosis, while the two recombinases, Rad51 and Dmc1, facilitate the recombination between homologous chromosomes during meiosis [54].
Nima-related kinases (Nek or NRK) represent a conserved family of serine/threonine kinases implicated in the regulation of distinct cellular events [47]. Neks have important roles in the maintenance of centrosome function and structure, mitotic microtubule organization, and the regulation of axonemal microtubule in cilia and flagella. Nima1 is an orthologue of Nek2 in humans which is involved in the maintenance of centrosome structures and mitotic microtubule organization, thus playing a role during mitosis. In Toxoplasma, Nima1 is essential for centrosome splitting, proper formation of daughter cell budding and faithful segregation of genetic material. A point mutation in a conserved portion of the gene causes a severe mitotic defect [48]. In Plasmodium, Nima1 is found in replicative forms of the parasites, in asexual and sexual stages, with a role in mitosis, and specifically in microgametes. In extracellular parasites, Nima1 showed almost double the expression levels in gamonts compared to parasites at 6 doc, and Nima2 and Nima4 were almost exclusively transcribed in gamonts/gametes (Fig. 5b). The mRNA levels in intracellular parasites found for Nima1 were significantly higher on 6 and 14 doc compared to 10 doc (Fig. 7 b, d). Our results correlate with the presence of Nima1 in asexual stages and sexual stages [49]. Due to the simultaneous presence of both in cell culture an upregulation in sexual stages could not be observed. By contrast, Nima2 and Nima4 were practically absent before 9 doc when the first mature gamonts occurred and peaked at 13 respectively 14 doc (Fig. 7b, e, f). In Plasmodium, Nima2 and Nima4 are only found in female gametocytes and the two encoded proteins are necessary for completion of the sexual cycle. In Nek2 knockout parasites premeiotic DNA replication is dysregulated and the parasites do not develop ookinetes, suggesting that the principle role of Nek2 is during DNA replication preceding the meiosis Nek 4 does not appear to be required for gametocytogenesis but is essential for premeiotic DNA replication in the zygote, consistent with the cell cycle related function [55]. As these two Nima genes were expressed in parallel with the occurrence of gamonts and unsporulated oocysts in vitro we assume that the encoded proteins have similar roles during the development of C. suis.
{kind=link}