Production of fatty alcohols by E. coli cells containing ADO
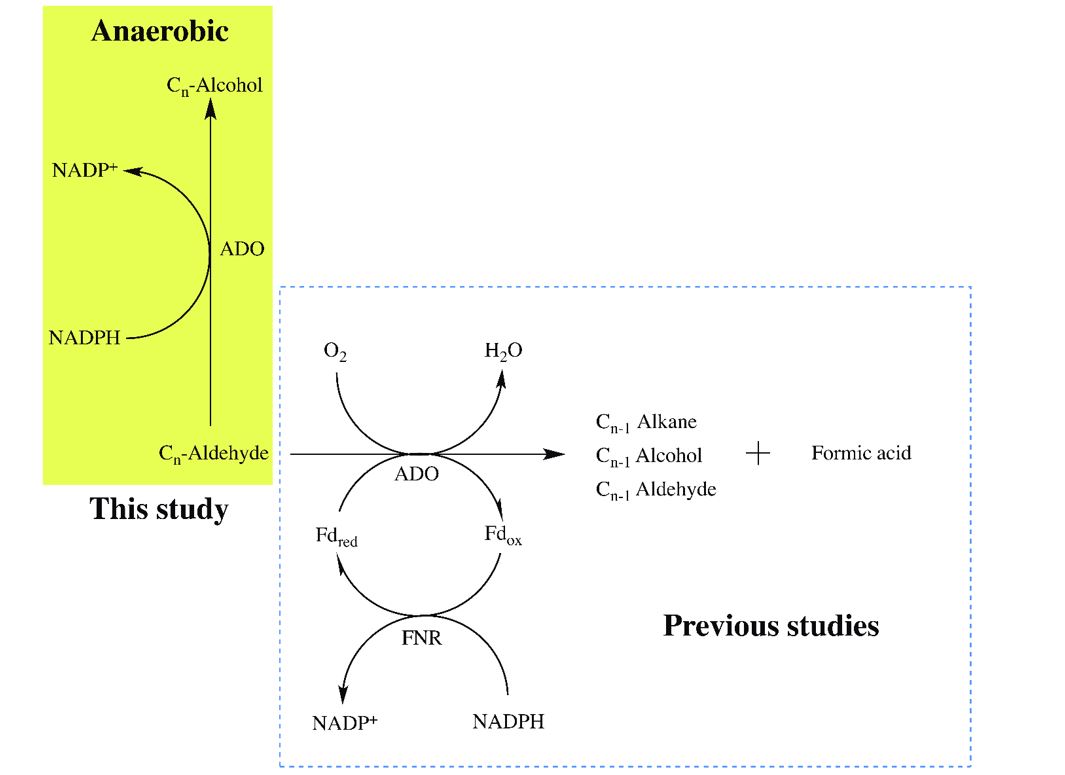
As illustrated in the previous report, alkane production can be produced from the synthetic metabolic pathway in Escherichia coli having aldehyde deformylating oxygenase (ADO) from Prochlorococcus marinus MIT9313, fatty acyl-CoA reductase (ACR1) from Acinetobacter baumannii and associated with redox partners ferredoxin (Fd) from Synechocystis sp. PCC 6803, ferredoxin (flavodoxin): NADP+ oxidoreductase (FNR). It was shown that addition of formate dehydrogenase (FDH) from Xanthobacter sp. 91 could increase intracellular NADH/NAD+ ratio in cells which can increase higher level of reduced Fd and increase yield of alkane production ~50% (23). Here, we further explored why the alkane product was limited at ~50% by further exploring whether this cell can generate other types of by-products. Analysis of metabolite profiles of the engineered cells containing PmADO and various types of auxiliary systems indicated that these cells produced significant amount of fatty alcohol, up to 25% fatty alcohols (Figure 1). This result prompted us to explore in-depth reaction mechanisms of fatty alcohol formation by PmADO using the purified enzyme and reconstituted systems.
Fatty alcohol production by the purified PmADO
Based on the results above showing that the metabolic engineered cell overexpressing PmADO could produce significant amount of fatty alcohol, we thus explored aldehyde reductase activity of the purified PmADO and also carried out experiments to rule out possible involvement of contaminating aldehyde reductases from the E. coli cell. As shown in Figure 2, the PmADO (with a subunit molecular weight of 24 kDa) obtained from our purification process (Experimental Procedures) is highly pure (>98% purity) with no other protein contamination visible by SDS-PAGE analysis. Based on the E. coli genome sequence, we identified putative aldehyde reductases produced in cells to monitor whether they could introduce possible false reductase activity in our PmADO assays. The analysis summarized in Additional file 1: Table S1 indicates that other genes identified as reductases (yahK, frmA, adhE, adhP, eutG, yqhD, yiaY and yjgB) all have their protein subunit sizes around 35-96 kDa. Results in Figure 2 clearly show that the purified PmADO obtained from our preparation has no (not even trace amount) contaminating protein in that region, ruling out a possibility of having other aldehyde reductases present in our enzyme sample. Using assays described in Experimental Procedures and in Table 1, the results indicated that the purified PmADO could catalyze fatty alcohol production from fatty aldehyde using various reducing systems (more results discussed below).
Fatty alcohol production by the unusual reaction of PmADO reductase activity
We first figured out nature of reductant required for PmADO aldehyde reductase activity and the effect of oxygen on this reaction. Reaction mixtures containing dodecanal and various %O2 (<0.0004%, 10%, and 15%) in the presence or absence of a reducing system, which are ferredoxin and ferredoxin reductase (Fd/FNR) were analyzed by GC/MS. Amount of fatty acid, alkane and alcohol products produced under various concentrations of oxygen and the reducing systems are shown in Table 1. The alcohol production by PmADO decreased when the O2 concentration was increased. The highest alcohol production was observed under anaerobic conditions with less than 0.0004% oxygen for both systems of PmADO with NADPH and PmADO with NADPH and Fd/FNR (Table 1). We noted that the alcohol production was not observed in the reactions without NADPH (Additional file 1: Figure S1).
Interestingly, dodecanal could be converted to dodecanol by PmADO in the absence of ferredoxin and ferredoxin reductase. The results showed that alcohol production yield by PmADO with NADPH (69 ± 5 µM) exhibited a similar value of product to the reaction of PmADO with the Fd/FNR reducing system (75 ± 4 µM) at less than 0.0004% O2 (Table 1). In the presence of the reducing system (Fd/FNR), the PmADO reaction showed an increase in alkane along with acid production when oxygen was increased (Table 1). However, the reaction of PmADO without the ferredoxin and ferredoxin reductase showed only acid in addition to the alcohol product (Table 1). These results indicate that the Fd and FNR are not necessary for transferring electrons from NADPH to produce alcohol by PmADO. Notably, PmADO can convert aldehydes to n-alcohols in the presence of only NADPH. As the highest yield could be obtained under anaerobic conditions, the data suggest that O2 is not necessary for the production of fatty alcohol by PmADO. Therefore, a minimum system for production of alcohol by PmADO only requires aldehyde and NADPH substrates (Scheme 1). Factors affecting production of fatty alcohol from aldehyde by PmADO including reducing systems and O2 concentrations are summarized in Table 1 and Figure 3.
To further address the effects of oxygen on production of fatty alcohol and to fine tune conditions for maximum alcohol production, reactions were performed in an anaerobic glove box and pre-incubated with an oxygen scavenging system glucose and glucose oxidase (Glc/GOx) to completely remove oxygen. Under these conditions and using octanal as a substrate, the detected octanol (80% conversion) was 20% greater than that of the reaction without oxygen scavenging system (60% conversion) (Figure 3B). Accordingly, no heptane or octanoic acid could be detected in these reaction mixtures. These data solidly demonstrate the conversion of fatty aldehyde to yield 80% fatty alcohol by aldehyde reductase activity of PmADO using NADPH as an electron donor in the absence of O2 and other redox partners.
To explore specificity of reducing equivalent for aldehyde reductase activity of PmADO, we investigated whether the enzyme can use NADH in addition to NADPH. The result showed that the PmADO aldehyde reductase activity could only use NADPH not NADH to produce fatty alcohol (Figure 4). Even with increased concentrations of PmADO (20, 40, 80, and 160 µM) in the presence of NADH, none of the reactions could produce fatty alcohols (Additional file 1: Figure S2).
The above findings showing that the purified PmADO could only use NADPH as a reductant also ruled out a possibility that the aldehyde reductase activity observed in this report comes from contamination of other aldehyde reductases produced in E. coli cells. Most of aldo–keto reductases in E. coli are known to use both NADPH and NADH as reductants to reduce a variety of aldehydes and carbonyl moieties [34, 46-49] (Additional file 1: Table S1). The only aldehyde reductase in E. coli which can use NADPH as a reductant is yqhD (Additional file 1: Table S1). As a subunit molecular weight of yqhD is 42 kDa, this indicates that the purified PmADO with a subunit molecular weight of 24 kDa (Figure 2) has no contamination from yqhD or other aldehyde reductases from E. coli. Altogether, all data suggest that the aldehyde reduction by NADPH was indeed catalyzed by the purified PmADO.
A ferrous (Fe2+) ion is not necessary for the aldehyde reductase activity of PmADO
The role of metal cofactor in aldehyde reductase activity of PmADO was investigated. We explored the involvement of Fe2+-cofactor in this activity by preparing an apoenzyme form of PmADO and measured its activity in comparison with Fe2+-bound holoenzyme. Although the native O2-dependent ADO reaction is involved with formation of an iron (III)-peroxo intermediate [34, 50] ,the aldehyde reductase activity to generate fatty alcohol is not involved with the metal cofactor and may not require the diiron-cofactor. Results from comparison of activities of ferrous supplemented PmADO, holo-PmADO and apo-PmADO (Figure 5) showed that the yield of fatty alcohol production from holo-PmADO (~60%) was similar to that observed in the reaction of apo-PmADO (~55%) and adding extra Fe2+ to the reaction (~75%). Based on standard variations of measurements, these values are all in the same range. Here, the results indicate that the metal cofactor is not involved in the reductase activity of PmADO. The MD simulations results (discussed later) also confirmed this conclusion.
The reductase activity of PmADO is highly specific towards medium- to long-chain aldehydes
Specificity of aldehydes that can be reduced by PmADO was investigated by carrying out the reactions of PmADO with medium- and long-chain (C6, C8, C10, C12, and C14) aldehydes and analyzed for alcohol production (Table 2, Figure 6). Based on kinetic parameters, the results indicate that PmADO does not exhibit strong chain length specificity with aldehyde substrates. This feature is consistent with specificity of aldehyde usage in the native PmADO activity in which both native cyanobacteria and reconstituted PmADO systems in E. coli can generate a wide range of alkane products [25, 51]. The results show that conversion of decanal to decanol gave the highest yield (98%) among all substrates used. Percentages of other aldehyde conversion to alcohol products were 86%, 79%, 68%, and 70% for hexanal, octanal, dodecanal, and tetradecanal, respectively. kcat and Km values of hexanal, octanal, and decanal are all relatively similar with a moderate trend of decreasing Km with increasing chain length of aldehyde from C6 to C10 (10.8 µM for C6, 10.3 µM for C8, and 6.5 µM for C10 (Table 2)). On the contrary, for the case of C12 and C14, the Km value increases with increasing chain length (Table 2); kcat of C12 is higher than that of C14. Altogether, these trends contribute to an approximately 3- to 5-fold changes in reaction efficiency (kcat/Km) from 15 × 10-3 μM−1min−1 for C10 to 3 × 10-3 μM−1min−1 for C12 and 5 × 10-3 μM−1min−1 for C14 (Table 2). It is important to note that no alcohol production was observed when 2-octenal was used as a substrate (Additional file 1: Figure S3).
Molecular docking and molecular dynamics (MD) simulations to identify a putative NADPH binding site
In order to identify a putative NADPH binding site in PmADO, MD simulations was used to dock NADPH and aldehyde into the PmADO active site. As decanal gave the highest catalytic efficiency in alcohol production by PmADO (Table 2), decanal was chosen to use in molecular docking and MD simulations studies. A structure of decanal was generated based on the structure of 11-[2-(2-ethoxyethoxy) ethoxy undecanal, which could be co-crystallized as a ligand in the PmADO crystal structure (PDB code 4PGK). NADPH was placed into the system containing PmADO and decanal. MD simulations were used to release stain of the protein and explore NADPH binding at the active site with the lowest energy point (-8.4 kcal/mol). The results showed that NADPH could bind to helices 1, 2, 4, and 5, which are close to the aldehyde binding site of PmADO. Snapshots taken around 0.54 ns of MD simulations are shown in Figure 7A. Moreover, the MD simulations also indicated that distances between Cα of Glu73 and Cα of Glu157 changed over the course of MD simulations, representing open and close forms of the PmADO enzyme. The open form could be observed before 0.54 ns (Cα of Glu73 and Cα of Glu157 bond distance increased from 12 to 20 Å) while the semi-closed form occurred after 0.54 ns (Cα of Glu73 and Cα of Glu157 bond distance decreased from 20 to 14 Å) (Figure 7B). These dynamics may represent the PmADO:NADPH:aldehyde complex conformational change prior to the next reduction step.
Additionally, the MD simulations of the PmADO:NADPH: decanal complex identified the distance between the C4-position of hydrogen atom of NADPH and the carbonyl carbon of fatty aldehyde (Figure 7A) as 3.8 Å. With this binding mode, the enzyme-bound NADPH can make stable contacts with decanal in the binding site of the PmADO. It should be mentioned that the PmADO structure used in our MD simulations (PDB code: 4PGK) was solved under aerobic conditions which may not represent the structure relevant to aldehyde reductase activity reported here. A hydride transfer distance in this structure may not be close enough for aldehyde reduction to occur. Up to now, no anaerobic crystal structure of an ADO enzyme is available. In the presence of molecular oxygen, the active site might be blocked, and consequently prohibiting the reaction between NADPH and aldehyde. In the case of aldehyde reductase, it is known that the hydride transfer from a nicotinamide co-substrate occurs only with a dehydrated aldehyde [52]. As aldehyde in solution likely exists in the hydrated form, thus during binding of a substrate, water must be stripped away for the substrate to prompt the compound for the catalysis [53]. This phenomenon explains the superiority of anaerobic alcohol production by ADO enzyme because molecular oxygen was removed for a proper binding of the enzyme substrate complex. The unique activity of PmADO that directly reduces fatty aldehyde by NADPH under anaerobic conditions provides structural and functional insights of PmADO for future biotechnology applications.
The reductase activity of recombinant PmADO enhances alcohol production in the metabolically engineered cell
To demonstrate the fatty alcohol production from reconstituted PmADO in E. coli under limited oxygen concentration, the PmADO and carboxylic acid reductase from Mycobacterium marinum (MmCAR) were constructed and measured fatty alcohol production in the metabolic engineered cell. MmCAR can catalyze fatty acid to aldehyde. Then, PmADO can use fatty aldehyde as a substrate to produce fatty alcohol. In order to avoid complications, decanoic acid was used as a supplement. The plasmids consisting of only MmCAR were also constructed to investigate the native aldehyde reductase activity from endogenous aldehyde reductase (AHR) in E. coli. Both plasmid systems were expressed in E. coli and the production of alcohol was quantified by GC/MS after 6 hours of a bioconversion process under various oxygen concentrations (<0.0004, 5, 10, 15, 20%). The results showed that fatty alcohol production could be observed in both MmCAR and MmCAR+PmADO under all oxygen concentrations (Figure 8). However, fatty alcohol production in the cell harbouring MmCAR+PmADO showed much greater amount of product than that of MmCAR cell factory, approximately 2-fold under almost all oxygen concentrations. The exception was found for the condition with <0.0004% O2 because both cell types showed low alcohol production (22%). This might be due to the disruption of energy metabolism in E. coli cells under strict anaerobic conditions [54]. Under 5% oxygen, MmCAR+PmADO gave the highest alcohol yield of dodecanol which is about 75%. This finding demonstrates that PmADO has the ability to enhance alcohol production in a cell factory as well as in vitro.
{kind=link}