Owing to the significant ecological and physiological constraints of reproduction, temperate bat species usually produce just one offspring per year [6]. However, some bats, including N. noctula, are capable of bearing twin offspring in a single litter as they have developed a uterus bicornis with two uterine horns and two equally functioning ovaries [7]. As reproduction is the most expensive of life-history traits, both energetically and nutritionally, no more than 20% of females tend to give birth to twins under natural conditions [37], though this may increase up to 88% in captivity [28]. The proportion of twins in our study was also higher (35% of females), most likely due to the stable microclimatic conditions, a lack of predation risk and stable access to food. Maximum fecundity, i.e. delivery of twins, is dependent on the condition of females in the early post-hibernation period and the maintenance of condition during pregnancy. Any increase in current reproductive output (maximum fecundity) is likely to occur only at the cost of future survivorship or future reproductive output, i.e. females are faced with the decision of investing in twins (actual reproduction success) or in single offspring (potential higher reproduction success in the future).
Female temperate bat species delay fertilisation until after hibernation [30, 38, 39] and must emerge in order to initiate pregnancy in early spring, when the weather may still be cold and food unavailable [40]. A female’s decision about actual reproduction strategy will then depend primarily on her fat reserves (perceived as body weight), as this influences both the amount of energy she can invest into embryo development, and the concentration of progesterone, the hormone that allows pregnancy to be maintained. In such cases, optimal conditions for successful reproduction will be ensured by utilising a hibernation strategy that maintains maximum fat reserves (cf. the “Thrifty Female Hypothesis”, [41] and through optimal timing of emergence from hibernation. Adult M. lucifugus females, for example, enter hibernation with higher fat reserves and spend these reserves more slowly than males during winter, presumably so that they can emerge from hibernation in good condition and initiate pregnancy [42]. Our heaviest females gave birth to twins regardless of length of hibernation and maximised their actual reproductive success. At the same time, we found that birth of twins was not observed in heavier but smaller females as a consequence of their longer hibernation (treatment group with later emergence). Births late in the year led to reduced skeletal growth, high juvenile mortality and poor long-term survival potential; thus, birth timing is a crucial factor in the mechanism as it controls the time available for growth before unfavourable feeding conditions develop in Autumn [43, 44]. While female M. lucifugus emerge earlier than males, females in the best condition emerge first, possibly in order to exploit more variable but potentially warmer microclimates at maternity roosts and occasional warm spring nights for foraging [45]. These factors could expedite gestation and support juvenile survival [25, 46].
In our study, it is likely that the lighter females with shorter hibernation (control group; short hibernation can be considered as the optimal emergence time due to the free availability of artificial food) resorbed one embryo in order to reduce their actual reproduction costs. It is often the case that female mammals can terminate a reproductive attempt that has a low probability of success and threatens future reproduction. Cases of embryonal mortality in bats may occur due to poor physical condition, resulting in blocking of egg implantation, resorbing of embryos and abortion of foetuses [27]. When abortion is not associated with expulsion of the products of conception from the uterus, the ensuing pyometra may result in death of the female [47]. In the big brown bat Eptesicus fuscus, implantation of five embryos has been demonstrated, though none of the females studied gave birth to more than two young. In the same species, occurrence of a developing foetus in the right uterine horn and a resorbed embryo in the left horn has also been confirmed [48]. In N. noctula, maturation of the Graafian follicle occurs immediately after birth, ensuring that all yearling females are capable of reproducing [7] and a strategy of producing one juvenile only (Strategy 1) appears to be beneficial for such young mothers. Nevertheless, termination of gestation remains somewhat debatable in wild animals as stress and other external ecological conditions, such as food deprivation, low food quality, high population density and/or genetic defects, can also play an important role [4].
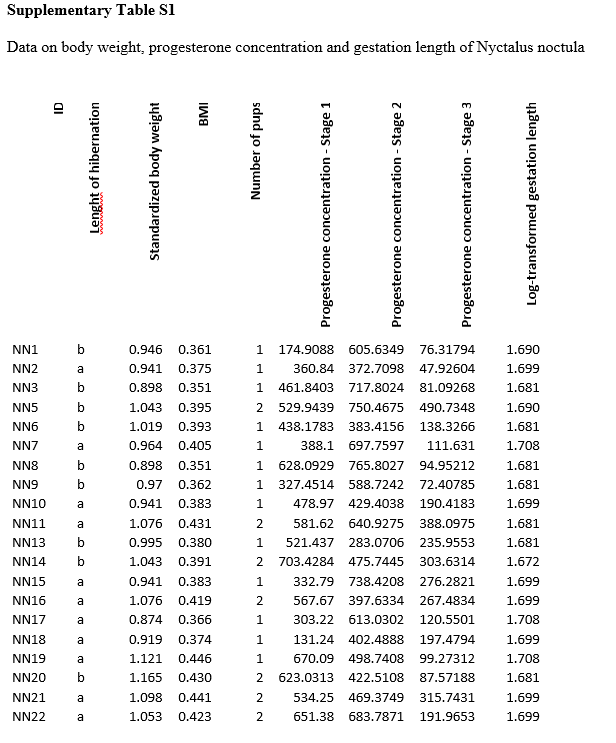
Fat reserves are important as the basic building block for production of progesterone, the hormone that maintains gestation [38]. In some mammals, the amount of progesterone in the blood is known to reflect the number of embryos [49]. We found that progesterone concentration in the early phase of gestation (Stage 1) was positively correlated with post-hibernation body weight. We were also able to describe two different patterns of progesterone development during gestation, enabling us to differentiate different reproduction strategies, i.e. females pregnant with single young, twins or single young after assumed embryo resorption. On the other hand, maximum concentration of blood progesterone (between 500 and 700 ng/ml) did not differ between the three strategies, suggesting that this represents the physiological value necessary for successful gestation. High progesterone concentrations in tree-dwelling N. noctula may reflect a need of a stronger physiological protection against embryo abortion due to the high impact of changing external conditions during gestation. Progesterone concentrations in M. lucifugus, which use buildings as summer maternity colony roosts [50], only reached up to 200 ng/ml during the later stages of gestation [32, 33], and progesterone concentrations are even lower in the polyoestrous tropical Arafura large-footed bat Myotis moluccarum, Australian C. gouldii or South African populations of M. schreibersii [34–36].
Our results showed that females that spent seven days longer in torpor (treatment group) gave birth to offspring an average of four days later than the control group. This suggests that they shortened the gestation period by two to three days in order to allow the juveniles to develop for longer during the optimal summer season with its rich food resources. In the greater horseshoe bat Rhinolophus ferrumequinum, pups born earlier are more likely to survive, increasing selection pressure to give birth to pups as early as possible [43]. This ability of female bats to manipulate the length of gestation has been known for some time [51], and a wide range of gestation lengths have now been recorded [13, 18, 28]. Indeed, our own estimation of 49-days, obtained under standard laboratory conditions with unlimited food and favourable temperatures, is similar to that calculated by [11]. The estimation of gestation length depends on two main factors, i.e. time of arousal from hibernation (as mentioned above) and subsequent environmental conditions. These factors are especially pronounced in temperate bat species as their offspring must grow quickly during the short warm season and accumulate resources to survive the winter [52]. On the other hand, it can be difficult to accurately determine time of natural ovulation in vespertilionid bats due to their use of sperm storage and delayed ovulation. In this study, we simply defined gestation length as the number of days between the end of hibernation (transfer to captivity) and the date of parturition (see [25, 53]). Unfortunately, there may be an element of bias in this estimation as we do not know the life history of the bats before they were transferred from their destroyed hibernation roost at the end of March. For example, the females may already have left the hibernaculum during warm nights and ovulation may have already occurred before the defined end of hibernation.
In bats, episodes of daily torpor under sub-optimal roosting conditions or periods of adverse weather will prolong gestation and delay parturition [25, 51], though the length of gestation is species specific and relatively stable in mammals [54]. The use of daily torpor and pregnancy are usually considered to be mutually exclusive processes, however their simultaneous occurrence is described in an increasing number of mammalian species [55]. In pregnant P. pipistrellus, it has been shown that the mean length of gestation is extended by induction of torpor at different stages during pregnancy, and that this extension is in good agreement with the period of torpor [51, 56]. Females will adopt such thermoregulatory behaviours during the gestation period in order to optimise juvenile development [26]. Under natural conditions, free-living reproductive bats tend to avoid torpor, especially long, deep bouts, while non-reproductive individuals do not [57]. Pregnant females may also fall into shallower and shorter bouts of torpor and maintain a higher body temperature [58, 59]. In our own study, where conditions were favourable (stable ambient temperature and food supply), there was no need for the females to utilise torpor at all. As such, we assume that the estimated gestation length is standard for N. noctula and that females optimise their behaviour to keep the length of the gestation as close to optimal as possible.




{kind=link}