Because of the increasing number of treatment failures and toxic effects by using current therapies for babesiosis, exploration and discovery of novel drugs with anti-Babesia activity is needed [3, 4, 22]. Babesia and Plasmodium share similar features in their biology as they are closely related genera among the apicomplexan parasites [23]. Due to this, some anti-Babesia drugs were initially investigated because of their antimalarial activity. Several showed excellent activities against Babesia, such as atovaquone combined with azithromycin for human babesiosis, and TAF, which was recently investigated for human and canine babesiosis [10, 24, 25].
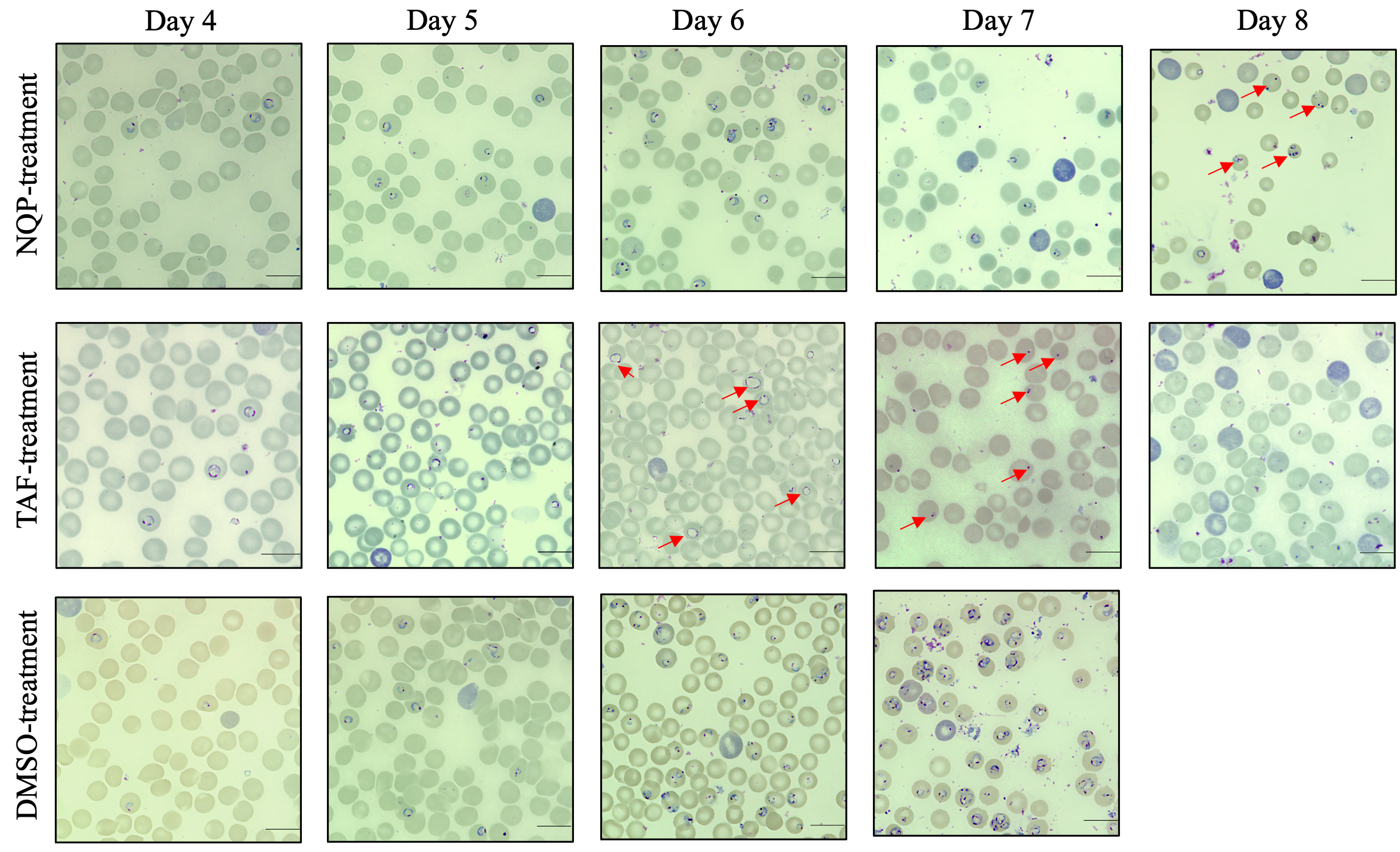
NQ is a 4-aminoquinoline antimalarial drug with a longer half-life but slower action than ART, and is currently combined with ART to treat malaria [14]. This combination therapy also inhibits Schistosoma mansoni [26]. Moreover, NQP combined with azithromycin for malaria treatment was also developed [27]. The toxic effects of NQP on mammalian host have been reported. Daily treatment in dogs for 14 days at a dose of 17.5 mg/kg/day and in rats for 70 mg/kg/day were considered safe. The safe doses in canine and rat models are equivalent to approximately 10 mg/kg/day in humans [15, 28]. A previous study reported that the concentration of NQP reached the peak level in plasma 2 hours post treatment with a dose of 10 mg/kg. The peak concentrations were 300.84 ng/mL and 273.29 ng/mL in plasma and erythrocytes, respectively, and the half-life of NQP was 198 hours (~8 days) in normal mice [29]. Interestingly, the concentrations were far greater in P. berghei-infected mice [29]. The anti-parasitic activity of NQP as demonstrated by the inhibition of B. gibsoni in vitro (Fig. S1) prompted us to further explore its anti-Babesia activity in vivo. We used the lethal species B. rodhaini in the mouse model for evaluating NQP as a therapeutic. In the current in vivo trial, NQP exhibited excellent inhibitory efficacy as evidenced by reduced parasite growth (Fig. 1a) and degenerative morphological changes in the parasites (Fig. 2). Furthermore, the first 2 days of treatment with 40 mg/kg NQP prevented the rise of B. rodhaini parasitemia starting from day 6 post infection compared to the typical rise of mean parasitemia in DMSO-treated mice. In addition, the accumulation of NQP in plasma [29] by completion of the 4-day treatment resulted in morphological changes of parasites in all treated mice at day 8 post infection. Moreover, the TAF-treated group showed an aberrant parasite phenotype (Fig. 2) which has been associated with oxidative stress [10]. B. rodhaini-infected mice in DMSO-treated group developed rapid anemia, whereas NQP and TAF prevented anemia development in infected mice (Fig. 1b).
Since TAF was approved by the U.S. Food and Drug Administration (FDA) as a single drug treatment for malaria, TAF studies have attracted much attention [10, 11]. Nonetheless, the limitation of TAF is the risk of inducing severe hemolytic anemia in individuals with G6PD deficiency in humans and the relapse of parasites, which are well documented [10, 24, 30]. Moreover, a single treatment of TAF on immunocompromised hosts could not eliminate parasites [10, 24]. Recently, TAF showed strong and broad anti-parasitic activity against Babesia spp., including B. microti, B. gibsoni, and B. rodhaini [10]. Hence, TAF was selected as a reference drug in this study. In the present study, the relapse of parasites was observed both in NQP-treated group and TAF-treated group (Fig. 1a). Therefore, NQP may need to be accompanied by other anti-Babesia drugs to augment its effect and prevent the regrowth of parasites.
In addition, the mechanism of action of NQP has not been fully elucidated. The inhibition activity of NQP for Plasmodium was hypothesized to be through the inhibition of hemozoin biocrystallization in the digestive vacuole of late-stage parasites and disruption of membrane system. Due to Babesia not producing hemozoin during parasite development, the inhibitory effect of NQP on the Babesia parasite is hypothesized to be related to targeting the parasite’s membrane system [15, 31].
It should be noted that there are some limitations to the present study. Although NQP exhibited a potential antibabesial effect, it has a slower onset of action and a longer half-life, which may easily lead to drug build-up with increasing the probability of developing resistance. Therefore, future studies are warranted to analyze the possible synergistic effect of NQP when administrated in combination with other drug which has a rapid onset of babesicidal action and a short half-life. Such analysis will help to determine the most effective composition ratio for treatment of Babesia in animals in clinical applications. Furthermore, the mode of action by which NQP inhibits the in vitro and in vivo growth of Babesia is still unknown. Subsequently, further studies are required to elucidate this point. Although the present study demonstrated the potential antibabesial efficacy of NQP in a mouse model, additional in vivo experiments are required to confirm such inhibitory effect in B. gibsoni- infected dog.
{kind=link}