Bread wheat is grown worldwide in temperate latitudes and subtropical regions (Royo et al. 2014) and constitutes the main source of proteins and calories in human diets (Venske et al. 2019). However, many wheat-growing areas are regularly exposed to heavy low temperature events during the early stage of wheat development (Braun and Sãulescu 2002) causing severe yield losses (Kajla et. al. 2015). Therefore, FroT is an important trait in breeding, in order to improve winter hardiness of wheat (Visioni et al. 2013).
In the recent years, high-throughput sequencing technologies fostered the availability of large SNP data sets and therefore the conduction of population genetic studies and GWAS (Wang et al. 2014). Furthermore, these technologies made the first fully annotated reference genome sequence of wheat available (IWGSC 2018). Altogether, this progress in plant genetics and genomics helps to increase the understanding of wheat biology and the molecular basis of important agronomic traits (Muthamilarasan and Prasad 2014; Li et al. 2020). Based on this progress, this study aimed to identify QTL regions and candidate genes associated with FroT in wheat.
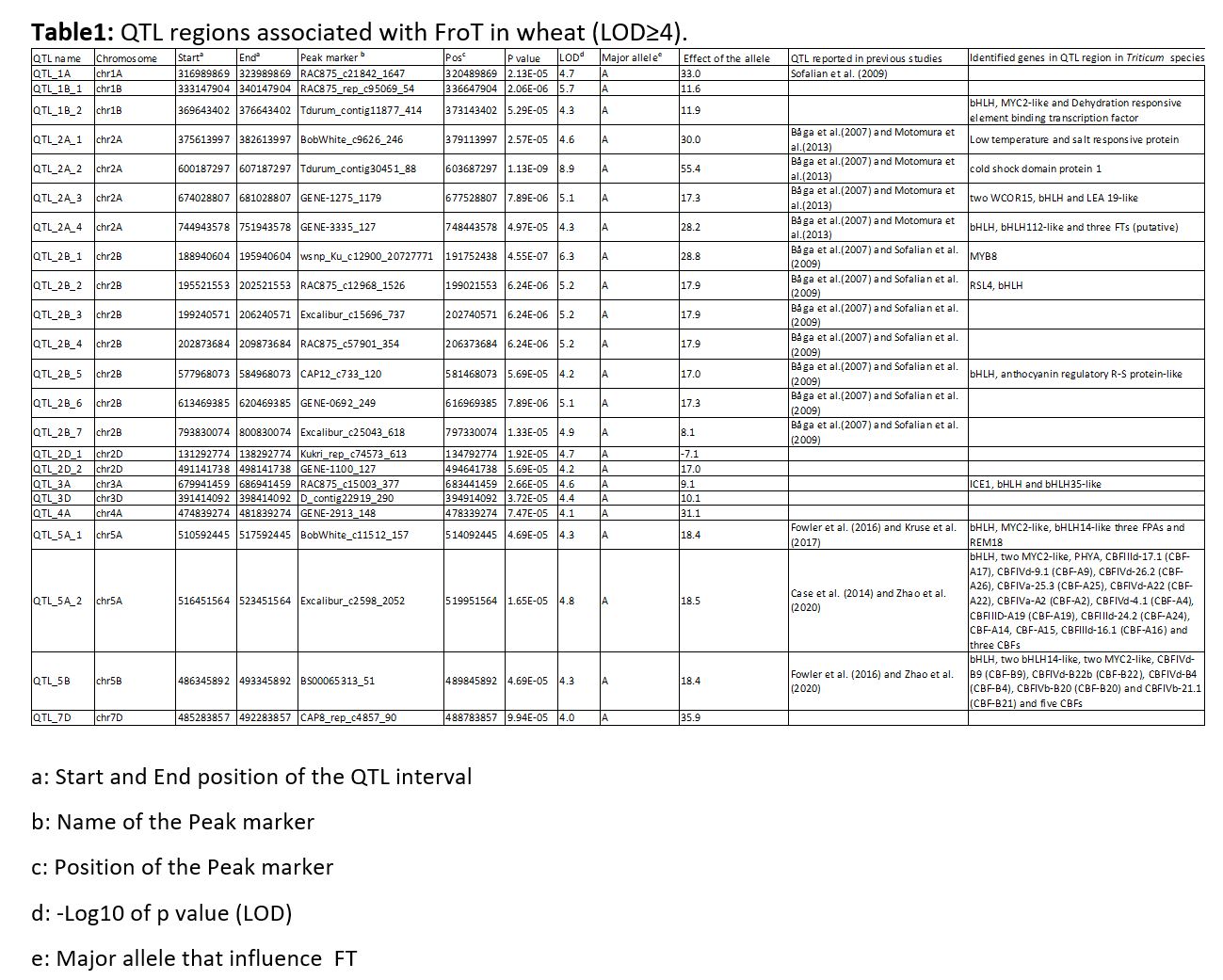
Recently, QTL mapping studies or GWAS identified several QTL regions associated with FroT in wheat on all wheat chromosomes except chromosome 4D (Zhao et al. 2020; Fowler et al. 2016; Zhao et al. 2013; Case et al. 2014; Kurse et al. 2017; Chen et al. 2019; Sieber et al. 2016; Babben et al. 2018). In general, it is difficult to compare QTL regions identified by different studies using different marker systems and different genetic maps. Therefore, to compare the records from literature with findings of this study, known flanking sequences of markers associated with QTL for FroT in literature were mapped to the reference genome sequence of Chinese Spring (IWGSC 2018). However, flanking markers were not available for all published QTL regions. Hence, these QTLs could not be anchored on the reference genome sequence of Chinese Spring. Furthermore, for some flanking marker sequences no unique position on the reference genome sequence of Chinese Spring (IWGSC 2018) could be identified. Therefore, for markers and QTL regions that could be not uniquely mapped to the reference genome, comparison with the results of this study was conducted based on the chromosome (supplementary Table S7).
To get comparable results, identified candidate genes in the current study with previous study (Babben et al. 2018), LOD≥ 4 was subjected as threshold for significantly associated markers with FroT. In addition, Zhao et al. (2020) reported associated significant markers with FroT with LOD≥ 4. The P value was also adjusted by Bonferroni-Holm (LOD≥ 5.5) correction in the present study. Finally, since only three markers were identified at Bonferroni-Holm threshold at LOD≥ 5.5 (result are not shown), we considered markers with LOD≥ 4 for further analysis. In total, 53 markers were found to be significantly associated (LOD≥ 4) with FroT in this study. These markers were assigned to 23 QTL regions on 11 chromosomes (1A, 1B, 2A, 2B, 2D, 3A, 3D, 4A, 5A, 5B and 7D).
In the present study, three out of 23 identified QTL regions namely QTL_5A_1, QTL_5A_2 and QTL_5B are overlapping with previously reported QTL regions (Case et al. 2014; Fowler et al. 2016; Kruse et al. 2017 and Zhao et al. 2020). It is known that important genes associated with FroT are located on chromosome 5A, e.g., CBF genes and VRN genes. In this study, 17 CBF genes, three bHLH family transcription factors and one FPA gene were identified within QTL regions associated with FroT on chromosome 5A and 5B. Polymorphisms within two out of 17 identified CBF genes (CBF-A14 and CBF-A15) were previously detected by CGAS (Babben et al. 2018). In total, 16 markers associated with polymorphic sites for CBF-A14 were significantly associated with FroT in the present study.
In plants, Hormones and photoreceptors, such as phytochromes, are regulated by CBFs (Kurepin et al. 2013). The positive role of PHYA on the transcription levels of CBF pathway genes is reported in tomato (Wang et al. 2020). In the present study, PHYA was associated with a QTL for FroT on chromosome 5A.
Furthermore, twelve QTL regions, i.e., QTL_1A, QTL_2A-1, QTL_2A-2, QTL_2A-3, QTL_2A-4, QTL_2B_1, QTL_2B_2, QTL_2B_3, QTL_2B_4, QTL_2B_5, QTL_2B_6 and QTL_2B_7 are potentially overlapping with previously reported QTLs on the same chromosomes. (Båga et al. 2007; Motomura et al. 2013 and Sofalian et al. 2009). However, due to the unavailability of flanking markers or due to the fact that the flanking sequences of available markers could not be anchored on the reference genome (IWGSC 2018); it was not possible to confirm this assumption. Three candidate genes associated with cold or low temperature tolerance were identified on chromosome 2A. The QTL_2A_1, QTL_2A_2 and QTL_2A_3 were co-localized with genes coding for Low temperature and salt responsive protein, cold shock domain protein 1, WCOR15 and LEA 19-like protein, respectively.
CORs are referred to as proteins encoded by cold-responsive or cold-regulated genes, which are involved in the cold tolerance acquisition and subsequent freezing tolerance. These genes, i.e., LEA, stress responsive protein (SRP), cold induced (KIN) and low temperature induced (LTI) (Guo et al. 2019), are increasing cold tolerance in plants. For instance, accumulation of COR/LEA proteins during cold acclimation protects cell structures and functions from freezing damage (Motomura el al. 2013). The Wcor15 is expressed under low temperature (Takumi et al. 2003) and encodes a chloroplast-targeted protein in wheat and barley.
QTL_2A_4 includes a gene coding for Flowering Locus T-like protein. It has been shown, that flowering time genes are not only responsible for the transition from the vegetative to the reproductive phase, but are also involved in various environmental stress responses. The relation between flowering and cold response is well known (Limin and Fowler. 2006).
For eight of the QTLs associated with FroT in this study, we did not find any evidence that these QTL were previously reported in literature. These QTLs are located on chromosome 1B, 2D, 3A, 3D, 4D and 7D and will be discussed in the following.
One out of two identified QTL regions on chromosome 1B contains a gene encoding a bHLH transcription factor. bHLH transcription factors play diverse roles in different physiological processes (Huang et al. 2013). Several studies have been shown that bHLH is involved in different responses, which are provoked by cold and other abiotic stresses in Arabidopsis and rice (Jiang et al. 2009; Kiribuchi et al. 2005; Ogo et al. 2006; Wang et al. 2003; Xu et al. 2014). Wang et al. (2019) identified 159 bHLH -encoding genes in wheat, which are involved in abiotic and biotic stress response. Furthermore, they pointed out that 98.7% of these genes are associated with more than one stress. In addition, the expression of these genes under different stresses was evaluated. In total, 38.44% of these genes were upregulated under cold stress in wheat (Wang et al. 2019).
The identified MYC2-like transcription factor is also called JAM (JASMONATE ASSOCIATED MYC2-LIKE) and bHLH14 is called JAM1. Both transcription factors are members of the IIId bHLH subfamily, which is phylogenetically closely related to MYC proteins interacting with JAZ proteins. These bHLH subfamily acts as transcription repressor of MYC2 and so as a negative regulator of jasmonate mediated response (Goossens et al. 2017; Sasaki-Sekimoto et al. 2013). Furthermore, Xiang et al. (2019) described the role of cold induced transcription factor bHLH112, which promotes a positive regulation of AP2/ERF transcription factor in Artemisia annua and Jiang et al. (2019) identified that bHLH35 is involved in cold tolerance in Anthurium andraeanum.
In addition, we identified one QTL region on chromosome 3A. This region comprises transcription factor ICE1 and bHLH transcription factor genes. The transcription factor ICE1 is known as an important gene involved in freezing tolerance (ICE -CBF -COR) pathway. ICE1 genes are known in wheat, but until now, no ICE1 gene was found to be located within a QTL region associated with FroT. Two ICE homologs, i.e., TaICE41 (accession no. EU562183) and TaICE87 (accession no. EU562184) have been identified in wheat (Guo et al. 2019). The identified AP2/B3-like transcriptional factor protein (REM18) on chromosome 5A is also a member of the DREB/ERF subfamily and it is accordingly maybe involved in FroT (Yamasaki et al. 2004; Chen et al. 2016).
The bHLH35-like gene was identified on chromosome 3A. Less knowledge is available for this gene. Jiang et al. (2019) have reported the positive role of bHLH35 in response to abiotic stresses in Arabidopsis. They reported that bHLH35 from Anthurium andraeanum (AabHLH35) increases stress tolerance to cold and drought in Arabidopsis. The expression of CBF1 and COR15A in wild type (WT) and AabHLH35 transgenic lines of Arabidopsis was significantly increased under cold stress compared to control plants. Expression of COR15A was 3-fold higher in AabHLH35 transgenic lines relative to WT lines. Therefore, they assumed that AabHLH35 might promote COR15A expression in response to cold stress. Furthermore, OsbHLH35 increased salinity tolerance in rice (Chen et al. 2018) and PebHLH35 from Populus euphratica increases drought tolerance in Arabidopsis (Dong et al. 2014).
As mentioned above, several genes are involved in the enhancement of FroT in plants. In addition to the ICE-CBF-COR pathway and flowering time genes, we identified nine genes of Jasmonates (JA) in the present study, which play a major role in the ICE-CBF-COR pathway (Figure 2) by activating transcription factors. Activated transcription factors bind to the cis-acting element in the promoter of target genes to increase FroT in plants (Ritonga and Chen, 2020).
{kind=link}