Pod pepper (Capsicum frutescens) is widely planted in China, especially around Wenshan city, Yunnan province, and viral diseases have now also become a major threat to pepper production in Yunnan. During July 2019, 89 pepper leaf samples were collected from three different fields in Wenshan. These samples were collected from Da Longshu village (a, 45 samples), Bai Shiyan village (b, 28 samples) and Da Shuduan village (c, 16 samples). All had typical viral symptoms of interveinal leaf yellowing and fruit discoloration (Fig. S1). These leaves (0.4 g per sample) were mixed into three pooled samples by origin, and then sent to the Central Laboratory of Zhejiang Academy of Agricultural Sciences (Hangzhou, China) for Next-generation RNA-Seq sequencing (NGS). A total amount of 1 μg RNA per sample was used as input material for the RNA sample preparations. RNA integrity was checked by Agilent 2100 Bioanalyzer (Agilent Technologies). The TruSeq RNA Sample Preparation Kit (Illumina) was used to construct cDNA libraries according to the manufacturer’s instructions. An Illumina NovaSeq 6000 platform with PE150 bp and CLC Genomics Workbench 20 (QIAGEN) was used for sequencing and data analysis. A total of 36,430,754 (a: 17,322,690, b: 11,735,694 and c: 7,372,370) paired-end reads were obtained, and 432,848 contigs (a: 143,689, b: 113,012 and c: 176,147) were generated de novo and compared with sequences in the GenBank nt using BLASTn, and 7 contigs were identified with E-values of zero. Contig_62 was 5992 nt long and had high identities (>87.5%) to the genome of PeVYV-3 (Pepper vein yellows virus 3) (KP326573;[3]) and the other contigs were matched with Chilli ringspot virus (ChiRSV, 3 contigs), Chilli veinal mottle virus (ChiVMV, 1 contig), Tomato zonate spot virus (TZSV, 1 contig) and Cucumber mosaic virus (CMV, 1 contig).
In order to verify the virus sequences, total RNA was isolated from each sample using TRIzolTM Reagent (Invitrogen) in compliance with the manufacturer’s instructions. Reverse transcription (RT) polymerase chain reaction (PCR) analyses were performed using the ReverTra AceTM qPCR RT Master Mix (Toyobo) and KOD-plus-Neo (Toyobo) following the manufacturer′ s protocol. RT was performed at 42°C for 60 min with random primers followed by 72°C for 10 min. The cycling conditions for the subsequent PCR were: 98°C 3min, and then 35 cycles of 98°C for 30s, 55°C for 30s, 68°C for 1kb/min; and 68°C for 10min. Primers designed from Contig_62 were used to amplify a coat protein (CP) fragment of the virus (PeVYV-CP) (Table. S1). Fragments of the expected size (621bp) were obtained in 58 of the 89 symptomatic samples but not from healthy plants (raised from seed in the laboratory). After adding dATP at the 3’- terminal of the PCR products using1 μL Taq™ (TaKaRa) at 72°C for 10min, they were cloned into pGEM-T Easy vector (Promega) and sequenced commercially (Sangon Biotech). The sequences were aligned using MUSCLE, and pairwise nucleotide sequence comparisons were done using the SDT (Species Demarcation Tool) v1.2 program[9, 10]. The 116 amplicons were 93.0-98.4% identical to PeVYV-3, and so we tentatively designated this virus in pod pepper as Pod Pepper vein yellows virus (PoPeVYV).
In order to acquire the full-length sequence of PoPeVYV in pod pepper, 5’ and 3’RACE reactions were then performed to obtain the complete terminal sequences. In brief, the 3’ end of the viral RNA was polyadenylated using Poly(A) Tailing Kit (Invitrogen) and first-strand cDNA was synthesized using M4T primer (Table. S1). The 3’ terminal end was PCR amplified from the cDNA using M4 and 3’-RACE-F (Tables. S1). Similarly, the 5’ end of the cDNA, after purification by treatment with Gel extraction Kit (Omega) was polyadenylated using the Poly(A) Tailing Kit. The purified cDNA/RNA heterocomplex was ligated with ZHM1 using T4 RNA ligase (Thermo Scientific) and PCR amplified using ZHM2 and 5’ RACE-R (Table. S1; Fig. 1A and B). To avoid errors in sequence assembly, the whole viral sequence was then amplified using two overlapping sections with the primer pairs PoPeVYV-1 and PoPeVYV-2 (Table. S1; Fig. 1C). RT-PCR products (expected sizes 4614 bp and 2824bp) were obtained using the methods of KOD-plus-Neo (Toyobo) and the two products (which overlapped by 1423 bp) were cloned into the pGEM-T Easy Vector (Promega) and sequenced. We obtained 14 amplicons of PoPeVYV-1 and 4 amplicons of PoPeVYV-2, and found that 12 amplicons of PoPeVYV-1 exactly matched the overlapping region of PoPeVYV-2. The complete sequence was 6015 nt long (GenBank accession number: MT188667).
PoPeVYV has a genome organization characteristic of members of the genus Polerovirus, with seven predicted genes encoding proteins P0 to P5 and P3a [6] (Fig. 1A). Over its entire genome our new virus is related (87.8% nucleotide identity) to Tobacco vein distorting virus (TVDV, accession EF529624) and had 85.0% nt identity to the (Chinese) PeVYV-3 (accession KP300822) (Table. 1). However, the 5’ half (nts 1-4251) of PoPeVYV has 93.1% nt identity to the corresponding region of PeVYV-3, and 87.7% nt identity to that of TVDV. The 3’ half (nts 4252-6015) of PoPeVYV has only 64.1% nt identity with PeVYV-3, but 91.7% nt identity with TVDV (Fig. S2). These results indicated that PoPeVYV in pod pepper might be a recombined virus with PeVYV-3 and TVDV as parents.
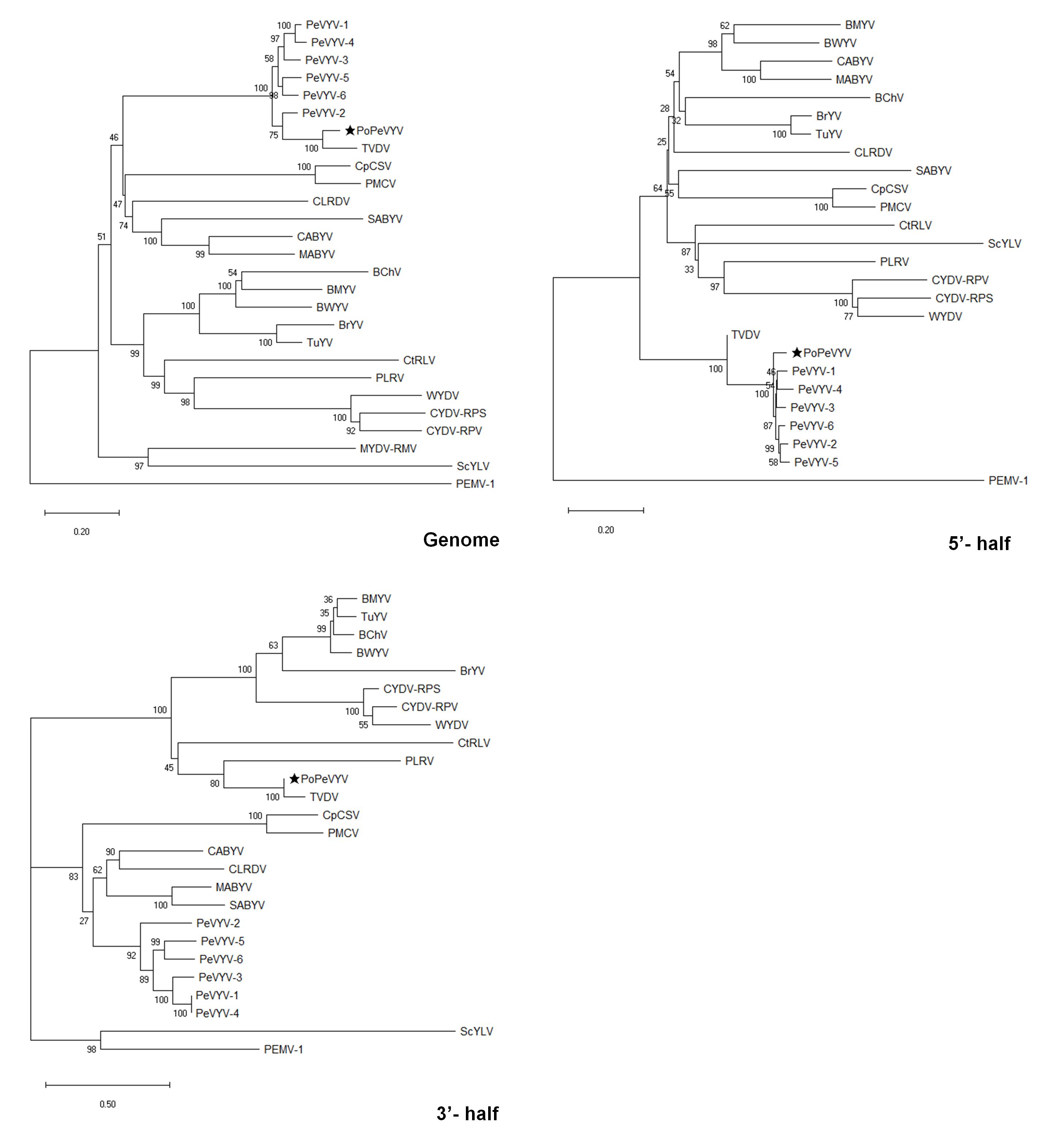
Poleroviruses are prone to recombination among themselves or with viruses belonging to other genera and the relationships between PoPeVYV, the previously described PeVYV isolates and TVDV suggests that there has been a recombination event affecting the 3’-end of the genome. This was confirmed using a variety of methods on the RDP4 recombinant platform [11]. A single recombination event affecting nucleotide positions 4126-5952, with TVDV and PeVYV-3 as the respective minor and major parents was consistently identified using GENECONV (P value of 1.035 E-93), RDP (8.410 E-85), BootScan (7.726 E-80), MaxChi (4.294 E-35), Chimera (7.719 E-05), SiScan (3.341 E-68), and Phylpro (2.331 E-15) (Fig. 2). Alignment of amino acid sequences of the PoPeVYV proteins with those of the six species of PeVYVs and TVDV also indicated a recombination event (Table. 1). Phylogenetic trees were constructed from the complete genome sequences, 5’- half or 3’- half of other poleroviruses and an enamovirus as an outgroup (Fig. S3). PoPeVYV, PeVYVs and TVDV formed an independent group separate from the other poleroviruses. The analysis confirmed that the genome and 3’ half of PoPeVYV was similar to TVDV, but that the 5’ half of PoPeVYV was similar to the PeVYVs (Fig. S3). This is reflected in the phylogenetic analysis of the amino acid sequences of the separate gene products [12]: the P0 of PoPeVYV is most similar to PeVYV-1/4, P1/P2/P3/P4 are closely related to other PeVYVs but the RTD is most similar to TVDV (Fig. 3). However, proteins encoded by the 5’ half of the genome (P0 to P4) had the highest identity to those of PeVYV-3 and PeVYV-6, whereas proteins translated from the 3’ half (P5/RTD) were more closely related to those of TVDV (Table. 1, Fig. 3). The various PeVYVs recognized usually have distinct P0 protein sequences and the topology of the P0 phylogenetic tree may depict the ongoing selection for a protein exhibiting better RNA silencing suppression capacity after a recent host jump [4, 7, 13]. The P0 of PoPeVYV has 77.5-87.1% amino acid identity and 85.2-90.5% nucleotide identity to that of the other PeVYVs, and 76.3% amino acid identity (81.3% nucleotide identity) to that of TVDV (Table. 1). In the P0 phylogenetic tree PoPeVYV was most similar to PeVYV-1 and PeVYV-4 and was clearly more similar to the PeVYVs than to TVDV (Fig. 3). These results suggest that PoPeVYV is a new recombinant polerovirus.
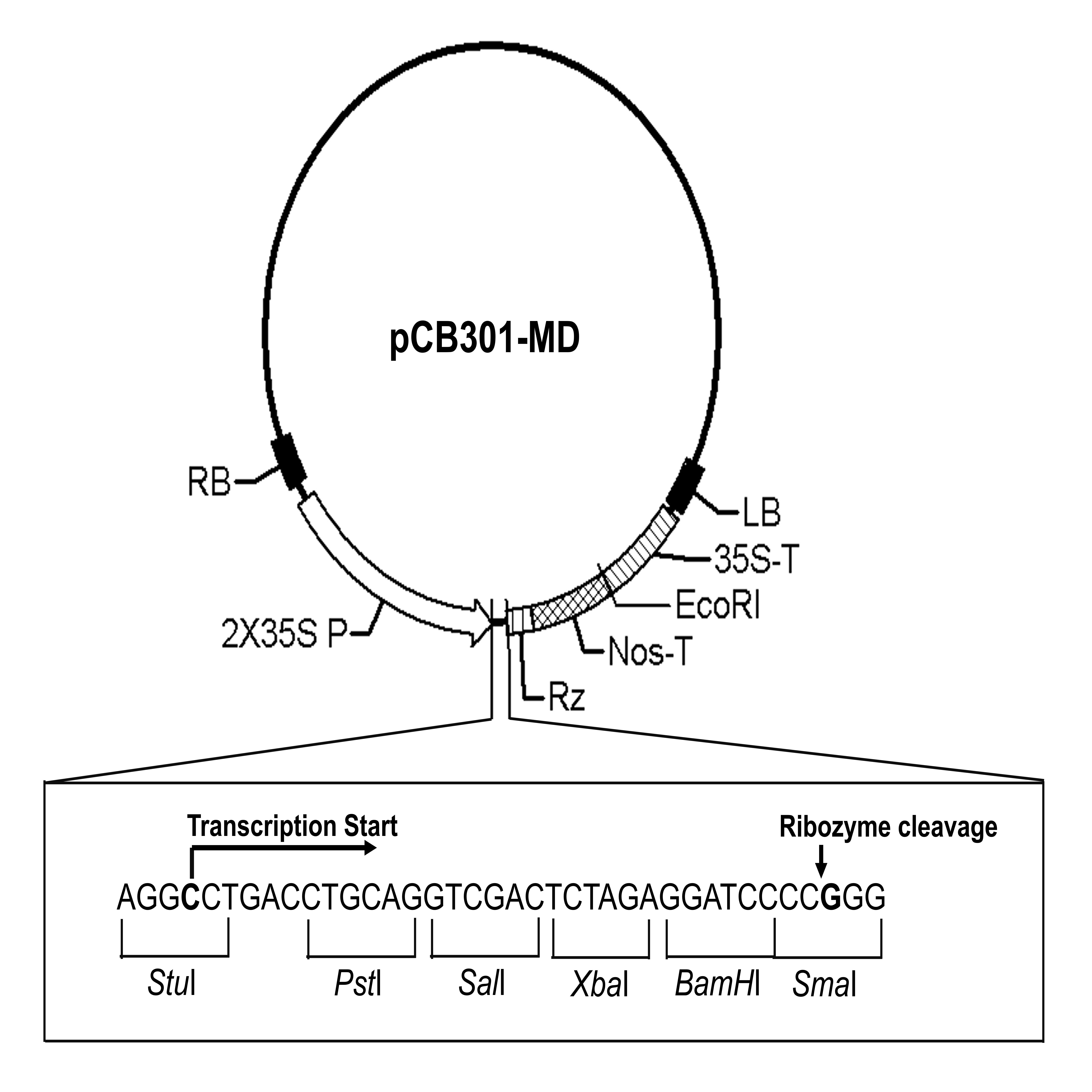
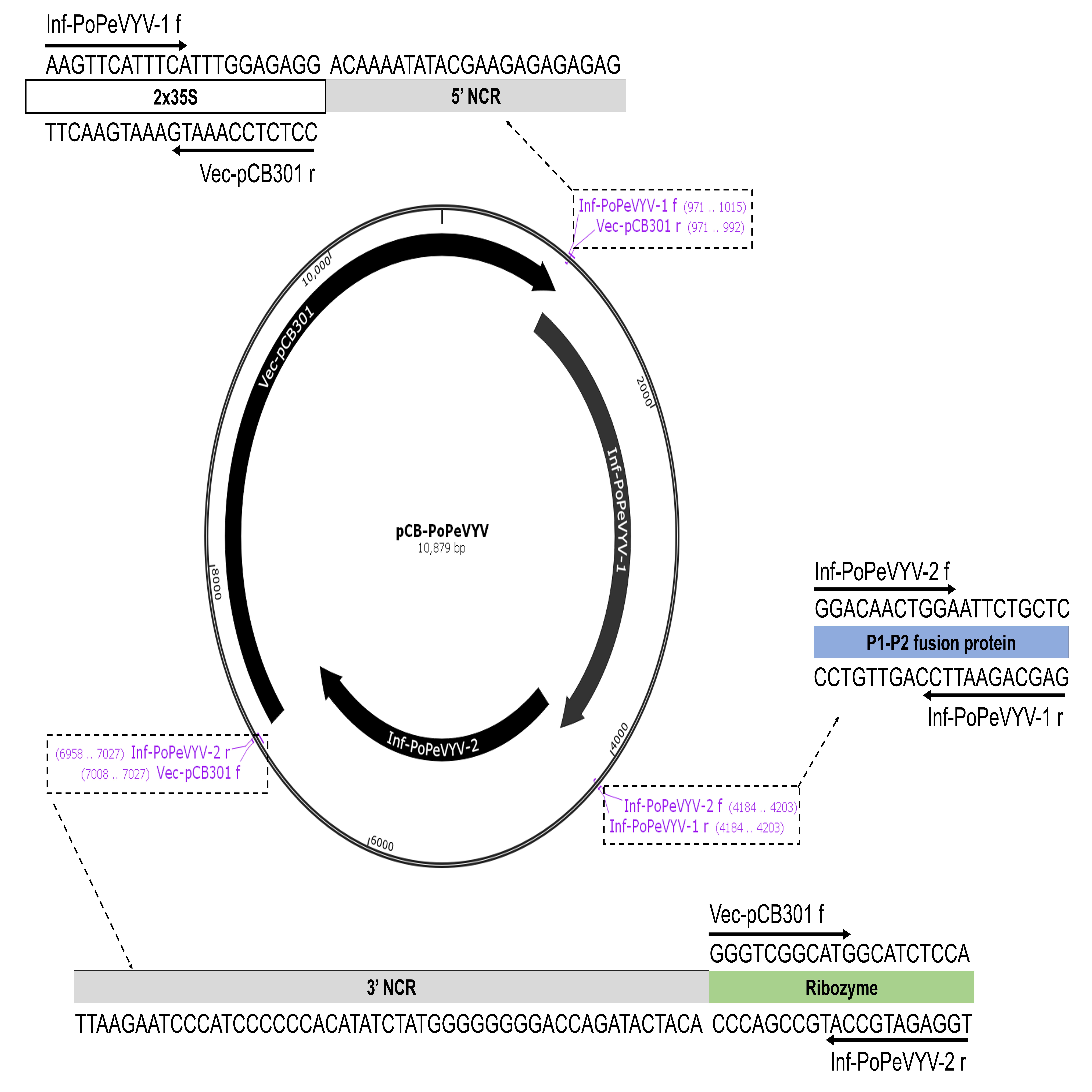
An infectious clone of the virus was constructed for further investigation. RT-PCR was performed using KOD-plus-Neo (Toyobo) following the manufacturer’s protocol. To generate infectious clones, the CloneExpress MultiS One Step Cloning Kit (Vazyme) was used for homologous recombination. Two overlapped PCR products were amplified with primers (Inf-PoPeVYV-1/2), and recombined with the linearized binary vector pCB301-MD (a modified version of pCB301 [14]), which includes the double 35S promoter and nopaline synthase terminator (NOS) (Fig.S4)[15, 16]. In this way, the full-length PoPeVYV cDNA was inserted between an upstream 35S promoter and a downstream hepatitis delta virus (HDV) ribozyme and NOS terminator in the binary vector to construct pCB-PoPeVYV. The PoPeVYV cDNA was ligated with 3’-terminal of 35S promoter without any extra nucleotide (Fig. S5)[15, 16]. This clone was transformed into Agrobacterium tumefaciens which was then delivered to C. frutescens plantlets by infiltration. There was mild upward leaf curling 45 days after inoculation (Fig. 4A), and RT-PCR using primers to detect the coat protein gene in the newly-emerged non-inoculated leaves showed that viral RNA was present and had spread systemically in all the inoculated plants (12/12) but not in the controls (Fig. 4B). Virions were purified from C. frutescens leaves using the method described previously [17]. Isometric particles about 25nm in diameter were observed in the purified preparation from the inoculated plants (Fig. 4C) but not from the controls. RT-PCR and subsequent sequencing confirmed that the virus in the symptomatic (systemic) leaves had the same sequence as the PoPeVYV cDNA clone inoculated (data not show). These results demonstrate the infectivity of the full-length PoPeVYV to C. frutescens.
In fields of cultivated pod pepper in Wenshan, we had observed severe yellowing and curling symptoms with small leaves (Fig. S1). Those symptoms were more severe than we observed in plants inoculated with the infectious clone of PoPeVYV by Agrobacterium tumefaciens which also did not have the obvious vein yellowing usually caused by PeVYVs. Previous studies have shown that the P0 proteins of poleroviruses display variable RNA silencing suppression activity, and induce distinct symptoms [18, 19]. The P0 of PoPeVYV has only 77.5-87.1% amino acid identity to the other PeVYVs, which could account for the distinct symptoms but the field plants were also infected with other viruses (PeVYVs, ChiVMV, ChiRSV, CMV etc) as frequently happens in the field and which has also been reported in Taiwan and Thailand [20, 21]. The severe viral symptoms in the field may therefore be a synergistic effect of mixed infection.
Recombination is an important source of genetic variability in viruses, particularly for viruses possessing an RNA genome. PeVYVs have higher identities to TVDV in the 5’ half of the genome and are considered to be recombinants of TVDV and other poleroviruses [4, 7, 8]. The new recombinant identified here has higher identity to TVDV at the 3’ part of genome, indicating a different sort of recombinant event. The 5’ half of the PoPeVYV genome shares high identity with PeVYV-3, which was reported from Hunan province in China [3]. The 3’ half is much more distant from PeVYV-3 but is more homologous with that of TVDV. TVDV cause a devastating tobacco disease in many tobacco producing areas include Hunan, Guizhou and Yunnan provinces in China [22, 23]. Additionally, PeYVY has been reported to infect tobacco in Guizhou province suggesting that co-infection of tobacco might have provided an opportunity for recombination [24]. Recombination poses a problem for classification. The currently-recommended species demarcation criteria in the family Luteoviridae suggest that different species should have >10% difference in amino acid sequence identity in any gene product from their closest relative. The P0 and P5 proteins of PoPeVYV have respectively 12.9-23.7% and 10.3-70.5% difference in amino acid identity to those of PeVYV1-6 (Table. 1). If applied here, those criteria suggest that PoPeVYV could be representative of a distinct species.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}