“Adapted” mutants arise out of independent “loss of function” strains upon long-term
cultivation on stress
Apart from molecular biology techniques and next-generation sequencing approaches,
classical vegetative growth assays were used to characterize potential functions of
genes associated with osmoregulation to study the HOG pathway in M. oryzae. Therefore, we cultivated the previously generated lof mutants ΔMohog1, ΔMopbs2, ΔMossk2, ΔMossk1, ΔMoypd1, ΔMohik1 and ΔMosln1 (20) on different stress-inducing media to compare them with the wildtype strain.
Mutants with an inactivated HOG pathway are sensitive to osmotic stress and resistant
to the fungicide fludioxonil (20). We noticed in the course of these assays that individual
mycelium parts grew out of the sensitive lof mutants ΔMohog1, ΔMopbs2, ΔMossk2, ΔMossk1 and ΔMoypd1 after cultivation for at least four to six weeks under continuous salt stress (Fig.
2).
Fig. 2:Example of Magnaporthe oryzae “loss of function” (lof) strains ΔMohog1 and ΔMohog1(adapted) after long-term cultivation upon salt stress. The lof strain ΔMohog1 was grown on complete medium (CM) inclusive 1 M KCl at 26 °C. The ΔMohog1 mutant (dark brownish colony) was found to be highly sensitive towards 1 M KCl, whereas
the outgrowing ΔMohog1(adapted) strain (bright colony) was able to grow much faster.
We isolated these individual mycelium parts in order to separate them as pure cultures
ready for further investigations and named them ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted), ΔMossk1(adapted) and ΔMoypd1(adapted). We have not been able to get adapted strains from the mutants ΔMosln1 and ΔMohik1 in the conditions tested so far. That underpins the hypothesis from our previous studies
that the two-component hybrid histidine kinase (HK) MoHik1p and the HK MoSln1p can
partially take over the function from each other (23) and, thus, the selection pressure
maybe not sufficient for an adaptation event in ΔMosln1 and ΔMohik1. Furthermore, it was not possible for us to isolate adapted strains from HOG pathway-independent
osmosensitive Magnaporthe-mutant strains, i.e. ΔMostu1 (transcription factor in cAMP/PKA signaling pathway, ΔMogpd1 (glycerol-3-phosphate dehydrogenase) or ΔMoskn7 (response regulator protein). Of course, we checked in the adapted strains whether
the genes originally inactivated in the “parent” lof mutants were still inactivated
in order to avoid any possibility of contaminations or confusions about mixed cultures.
We did this for all adapted strains using ITS-sequencing and southern blot analyses
(supplementary Fig. S2).
In empirical investigations, a combination of different vegetative growth assays was
used to characterize the “rapid adaptation frequency” in the different lof mutants.
Twenty mutant strains of each of ΔMohog1, ΔMopbs2, ΔMossk2, ΔMossk1, and ΔMoypd1 were grown continuously on solid complete medium (CM) including 1 M KCl or 1.5 M sorbitol
as stress-inducing agents. Several adapted strains arose out of the lof mutants in
each plate, being able to grow much faster under the stress condition. The adapted
strains were then transferred onto CM without stressors for the following two weeks.
Subsequently, we transferred the colonies onto repeated stress medium to investigate
in which strains the “adaptation” is stable and identified the stably adapted strains
(Fig. S3). Notably, we were able to identify more adapted strains from ΔMopbs2 and ΔMoypd1 under salt stress than under sorbitol stress. By contrast, we obtained more adapted
strains under sorbitol stress than under salt stress from ΔMossk2 and ΔMossk1 (Supplementary Fig. S3).
Osmoregulation is permanently restored in the adapted mutants
The strains ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted), ΔMossk1(adapted) and ΔMoypd1(adapted) displayed significant differences in growth speed on salt stress compared to their
“parent-strains,” the lof mutants ΔMohog1, ΔMopbs2, ΔMossk2, ΔMossk1 and ΔMoypd1 (Fig. 3 [1]). All lof mutants were strongly sensitive towards 1 M KCl stress, whereas
all the adapted strains were less sensitive, similar to the wildtype strain (Fig.
3 [1], lower row, colonies [A]). Particular attention has to be paid to the finding
that adapted strains, which were pre-cultivated on normal CM (unstressed conditions)
and then transferred back to the repeated salt stress, were found to grow as fast
as those taken directly from stress conditions and, thus, being able to adapt to KCl
stress immediately (Fig. 3 [1], colonies [C]). In conclusion, the mutations/alterations
within ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted), ΔMossk1(adapted) and ΔMoypd1(adapted) appear to be stable. Similar results to the growth assays on solid media could be
observed in liquid cultures upon KCl stress (Fig. 3 [2]) and sorbitol stress (supplementary
Fig. S1).
Fig. 3:
[1] Stable reestablishment of osmoregulation in adapted strains. Vegetative growth of the Magnaporthe oryzae wildtype strain, mutants with inactivated components of the HOG signaling cascade
and the “adapted” strains upon salt stress. The fungal colonies were grown on CM (upper row) and CM including 1 M KCl (lower row)
for 7 d at 26 °C. A, B and C are representative for each plate. A = lof mutant, B = “adapted” strain taken directly from CM inclusive 1 M KCl (“adaptation” conditions),
and C = “adapted” strain after six weeks of preincubation on CM (to have long-term unstressed conditions before this assay). In the case of the wildtype strain (left), the three
cultures on each test plate used in the assays were from the M. oryzae wildtype strain 70–15 cultivated in an equal manner.
[2] Mycelium dry weight of the Magnaporthe oryzae wildtype strain, mutants with inactivated components of the HOG signaling cascade
and the “adapted” strains after growth in liquid culture upon KCl stress. The fungal colonies were grown in 250 ml CM including 1 M KCl for 6 d at 26 °C and
120 rpm. Error bars represent the standard deviation of three biological replicates
of each strain.
Glycerol is the major compatible solute produced by the adapted strains after salt
shock
Intracellular production of compatible solutes was determined by high-performance
anion-exchange chromatography with pulsed amperometric detection (HPAEC-PAD) and compared to compatible solute production of the lof mutants and the
wildtype strain to further investigate the phenomenon of restored osmoregulation in
ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted), ΔMossk1(adapted) and ΔMoypd1(adapted). Hyperosmotic shock was imposed by 0.5 M KCl stress towards the fungal strains, and
the intracellular levels of the major osmolytes mannitol, trehalose, arabitol and
glycerol were determined. No increases in the mannitol and trehalose levels were detected
in the lof mutants and the adapted strains after osmotic shock (data not shown). A
slight increase in the mannitol level was detected and no significant change for the
trehalose level of the wildtype strain (data not shown). Similar to data from (19),
arabitol was found to be the major intracellular compatible solute produced by the
wildtype strain after osmotic shock (Fig. 4).
Fig. 4:Compatible solute production in the Magnaporthe oryzae wildtype strain (WT 70-15), the lof mutants and the “adapted” strains after osmotic
shock. Compatible solute accumulation (glycerol and arabitol) was assayed in mycelium after
7 h salt stress. The mycelium was grown for 72 h in CM (2 % glucose) before being
shocked with 0.5 M KCl for an additional 7 h. Carbohydrates were extracted and quantified
by liquid chromatography. Error bars represent the standard deviation of three biological
replicates of each strain.
By contrast, the lof mutants ΔMohog1, ΔMopbs2, ΔMossk2, ΔMossk1 and ΔMoypd1 were not able to produce either arabitol or glycerol in significant amounts. Interestingly,
it was found that all the adapted strains ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted), ΔMossk1(adapted) and ΔMoypd1(adapted) responded to hyperosmotic stress by accumulating high amounts of glycerol rather than
arabitol (Fig. 4). Based on these observations, we conclude that glycerol may somehow
compensate for the lack of arabitol upon salt stress in the adapted strains.
Fludioxonil susceptibility is restored in the adapted mutants
Vegetative growth assays were conducted using minimal medium (MM) and MM including
10 µg/ml fludioxonil to further investigate whether fludioxonil-susceptibility and
not only osmoregulation is restored in ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted), ΔMossk1(adapted) and ΔMoypd1(adapted) (Fig.5).
Fig. 5:
Stable vegetative growth of the Magnaporthe oryzae wildtype strain (WT 70-15), mutants with inactivated components of the HOG signaling
cascade and the “adapted” strains upon fludioxonil treatment. The fungal colonies were grown on minimal medium (MM, upper row) and on MM including
10 µg/ml fludioxonil (lower row) for 7 d at 26 °C. A, B and C are representative for
each plate. A = lof mutant, B = “adapted” strain taken directly from CM including 1 M KCl (“adaptation” conditions)
and C = “adapted” strain after six weeks of preincubation on CM (to have long-term unstressed conditions before this assay to show the memory effect). In the case of the wildtype
strain (left), the three cultures on each test plate used in the assays were from
the M. oryzae wildtype strain 70–15 cultivated in an equal manner.
Apart from the osmoregulation capacity, fludioxonil sensitivity was reconstituted
in the adapted strains. The HOG pathway lof mutants were resistant towards the fungicide
(Fig. 5, colonies [A], lower row), whereas all adapted strains were susceptible, but
not quite as strongly as the wildtype strain(Fig. 5, colonies [B], lower row). Similar to the findings concerning salt stress
presented previously in Fig. 3, the adapted strains, which were pre-cultivated in
unstressed conditions for a long time, showed the same phenotype as the adapted strains
taken directly from stress medium (Fig. 5, colonies [C], lower row).
Fludioxonil sensitivity in the adapted strains is not dependent on the production
of arabitol or glycerol
Fludioxonil induces a hyperactivation of the HOG pathway and, thus, a continuous stress
response towards high osmolarity. We found, by using HPAEC-PAD analytics, that fludioxonil
treatment resulted in the production of arabitol and glycerol in the wildtype strain
(Fig. 6). We conducted the experiments with the lof mutants and the adapted strains
ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted), ΔMossk1(adapted) and ΔMoypd1(adapted) to check whether fludioxonil-dependent production of the compatible solutes was altered
in the adapted strain. The fungal cultures were grown in liquid CM (2 % glucose) and
stressed with 10 µg/ml fludioxonil. As expected, we did not find increased compatible
solute production upon fludioxonil-treatment in the lof mutants. Exemplarily, we present
the data for ΔMohog1 (Fig. 6).
Fig. 6:Compatible solute production in the wildtype strain, ΔMohog1 and ΔMohog1(adapted) upon fludioxonil treatment. Compatible solute accumulation (glycerol and arabitol) was assayed in the mycelium
of the wildtype strain, ΔMohog1 and ΔMohog1(adapted) after 7 h treatment with 10 µg/ml fludioxonil. The mycelium was grown for 72 h in
CM (2 % glucose) before being shocked with 10 µg/ml fludioxonil for an additional
7 h. Carbohydrates were extracted and quantified by liquid chromatography. Error bars
represent the standard deviation of three biological replicates of each strain.
Interestingly, no increase of arabitol or glycerol production was detectable in the
fludioxonil-susceptible adapted strains in the presence of the fungicide (Fig. 6).
It has to be pointed out that the metabolic response of the adapted strains after
fludioxonil treatment is different compared to the compatible solute production we
observed upon KCl treatment (Fig. 4). All the adapted strains ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted), ΔMossk1(adapted) and ΔMoypd1(adapted) responded to salt stress by accumulating high amounts of glycerol, whereas this is
not the case under fludioxonil stress. In conclusion, fludioxonil sensitivity in the
adapted strains does not appear to be dependent on compatible solute production.
Reestablished osmoregulation does not complement reduced virulence of the lof mutants
The lof mutants of the HOG pathway in M. oryzae were found to be reduced in virulence towards rice plants compared to the wildtype
strain. Interestingly, ΔMohog1(adapted), ΔMopbs2(adapted), ΔMossk2(adapted) and ΔMossk1(adapted) were not found to be as virulent as the wildtype strain, and rather less virulent
than the lof mutants (Fig. S4). We were not able to conduct the pathogenicity assays
regarding ΔMoypd1(adapted), since the mutant failed to produce conidia exactly like ΔMoypd1 (20).
There are no relevant structural variations on DNA-level in the genomes of adapted
strains
We had a deeper look at the genomes of ΔMohog1 compared to ΔMohog1(adapted) in order to find out the cause of that rapid adaptation in the adapted strains. We
also added data from the genome sequencing of ΔMopbs2(adapted) to strengthen the analysis and narrow down the outcome of putative candidate genes
showing structural variations in the adapted strains. Single nucleotide variations
(SNVs) and short indels were detected for ΔMohog1(adapted), ΔMopbs2(adapted) and ΔMohog1 in comparison with the reference sequence of the wildtype strain. The resulting variants
were further annotated based on their chromosomal location and biological effects,
such as synonymous/non-synonymous single-nucleotide polymorphisms (SNPs), upstream/downstream,
untranslated regions (UTRs) and intergenic regions. The ratio of transition and transversion
was also calculated for single nucleotide variation. Over 80 % of all SNPs detected
were found to be located outside exons and a significant enrichment in regions adjacent
to exons and UTRs was detected (data not shown). Furthermore, in silico protein modelling suggested that several non-synonymous SNPs are probably direct
targets of selection, as they lead to amino acid replacements in functionally important
sites of proteins. Hence, the structural variation discovery analysis of small-scale
(< 20 bp) and large-scale variations (> 20 bp) such as frameshift, stop codon insertion
resulted in a list of three genes (five putative gene variants) in the overlap of
ΔMohog1(adapted) and ΔMopbs2(adapted), but none of them leads to a protein effect. The variations are only transitions
not changing the amino acid composition of the corresponding proteins. Furthermore,
we checked the homologous gene loci of the known yeast suppressor mutation genes RSG1 (RHB1) (24), SOO1 (25), SGD1 (26) and PMK1 (KSS1) (27) intensively. Within all these loci, we could not identify structural variations in
the genome of the adapted strains ΔMohog1(adapted) and ΔMopbs2(adapted).
Since regulatory gene elements, such as promoters, are of prior interest regarding
their direct influence on gene expression alteration, we decided to investigate the
presence of genetic structural variations in the putative promoter regions of all
annotated genes in the M. oryzae genome. The promoter region was defined as the region on the genomic DNA 1500 bp
upstream of each annotated gene start codon. We performed a structural variation discovery
analysis of small-scale (< 20 bp) and large-scale variations (> 20 bp) resulting again
in no significant variations in the overlap of the promoter regions of ΔMohog1(adapted) and ΔMopbs2(adapted) (Fig. S5 [A]). We considered here only insertions and deletions as a “polymorphism
type” and insertions, truncations and frame-shifts as a “protein effect.”
Differential transcriptomic profiles in the adapted strains in response to osmotic
stress
Next-generation sequencing analysis of RNA samples from the wildtype strain, ΔMohog1 and ΔMohog1(adapted) before and after 25 min salt stress (0.5 M KCl) should present insights into transcriptional
changes which may be responsible for the adaptation phenomenon observed (methods for
cultivation before RNA-isolation, see (28)). A principal component analysis was performed
to characterize the relationship between the strains analyzed. The processed transcriptome
data of ΔMohog1 (untreated), ΔMohog1 (25 min 0.5 M KCl-stress), ΔMohog1(adapted) (untreated) and ΔMohog1(adapted) (25 min 0.5 M KCl-stress) appears to form distinct clusters within each sample group
of strain samples investigated (Fig. 7A).
Fig. 7:
RNA-sequencing (Seq) data processing. (A) Principal component analysis results. The figure displays a three-dimensional scatter plot of the first three principal
components (PCs) of the data. Each point represents an RNA-Seq sample. Samples with
similar gene expression profiles are clustered together. Sample groups are indicated
by using different colors as indicated in the legend provided. (B) Heat map of transcriptional regulation patterns of the wildtype strain, ΔMohog1 and ΔMohog1(adapted) after 25 min salt stress (Clustergrammer analysis). The figure contains a heat map displaying the expression profile of the top 500
differential expression genes (DEGs) for each sample in the RNA-Seq dataset. The set
of mutant strains ΔMohog1 and ΔMohog1(adapted) prior and after salt stress was compared to wildtype strain 70-15 (indicated in the
figure as sample group) for perturbation contrast. Each row of the heat map represents
a gene, each column represents a sample and each cell displays normalized gene expression
values.
Since the principal component analysis revealed distinct differences on transcriptomes
of corresponding strains, we decided to follow up by clustering the genes according
to their expression followed by construction of a co-expression network. In the clustering
analysis, represented by a heat map visualization, the fact that the transcript value
is strongly different in ΔMohog1(adapted) compared to the WT 70-15 and ΔMohog1, even in untreated conditions, is clearly visible (Fig. 7 [B]). Cluster 6 is significantly
exclusively upregulated and cluster 1 is harboring exclusively down-regulated genes
in ΔMohog1(adapted), whereas exclusively up-regulated genes in the WT 70-15 could be found in cluster
2 (Fig. 7 [B]).
Glycerol metabolism-associated genes are affected in the adapted strains
To investigate, whether the groups of differentially expressed genes (DEGs) are functionally
related, we performed a gene ontology (GO) enrichment analysis to determine the biological
functions associated to them. Generally, this approach helps to highlight groups of
genes with coherent biological functions that are presumably acting in coordination
in response to salt stress. The clustering analysis was employed over the whole gene
co-expression network and in a selected subset or cluster of interest. The result
indicated that there were several significantly enriched terms of DEGs, from which
the most representative are “lipid biosynthetic process,” “glutamine family amino
acid metabolic process,” “carbohydrate transport,” “dephosphorylation” and “carboxylic
acid biosynthetic process” (Fig. 8).
Fig. 8:Network visualization and analysis obtained by the Cytoscape/ClueGO clustering pipeline. The size of the nodes reflects the statistical significance of each term. Subnetworks
of genes that are highly connected (network cluster) in a co-expression network are
usually involved in similar biological functions. The cluster analysis of the network
was performed to find groups of genes that may be acting in a coordinated manner. A
gene ontology (GO) enrichment analysis was conducted using ClueGO to determine the
biological functions associated to them to investigate if a group of genes are functionally
related. The name of each group is given by the most significant term of the group.
This analysis was performed over the whole network.
This enrichment analysis suggests that the highly connected genes from the largest
cluster in the network are involved in the regulation of complementary processes triggered
by salt stress.
In order to follow up the results of glycerol-production in the adapted strains, we
further investigated most of the genes potentially contributing to the production,
metabolism or transport of glycerol. However, we checked these genes presented and
added a list of homologous genes potentially related to the production, metabolism
or transport of glycerol from a database- and literature-based approach, respectively,
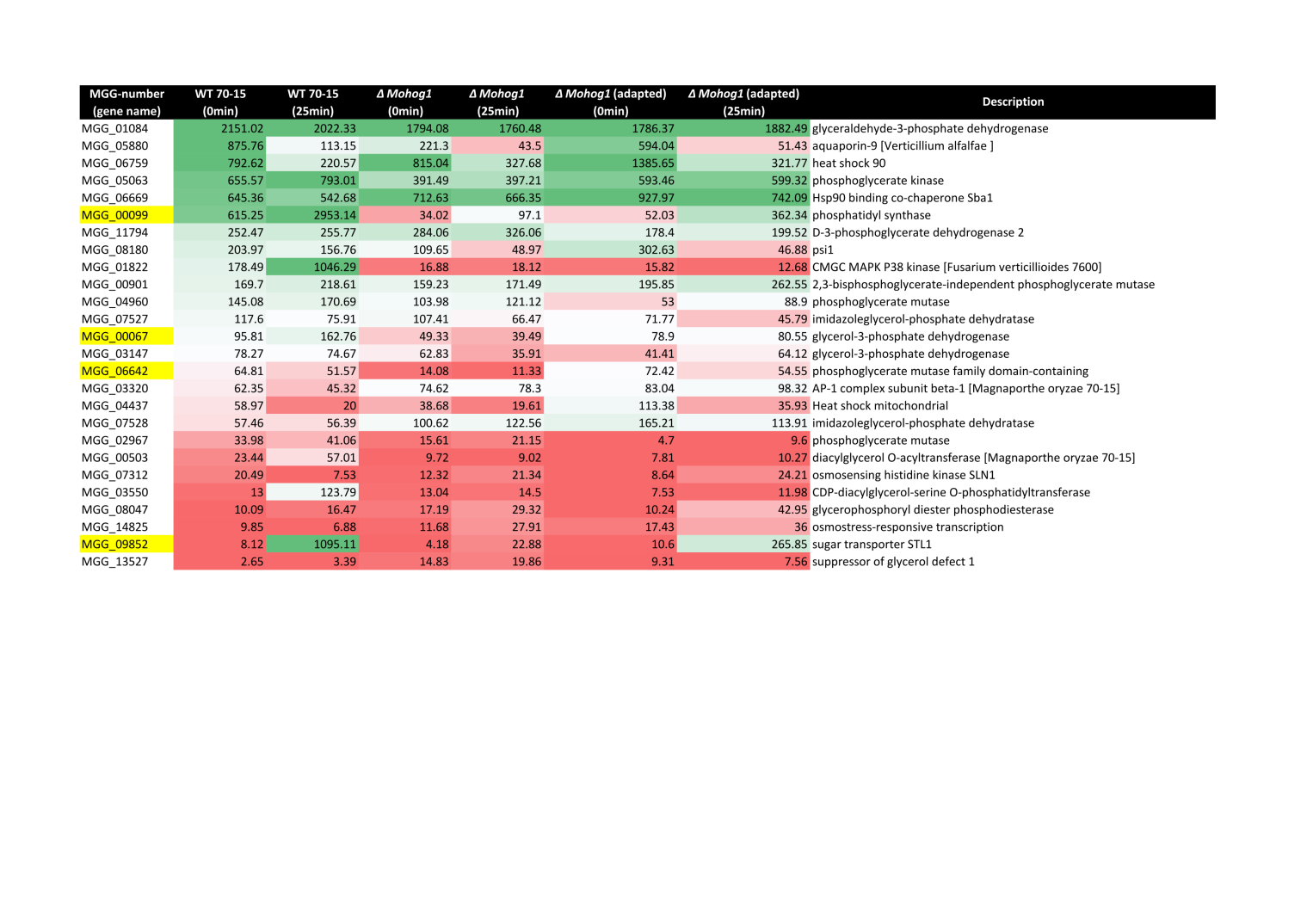
from knowledge of our own unpublished research activities (Tab. 1).
Tab. 1:Glycerol biosynthesis and osmotic stress response-related genes being differentially
expressed across the strains analyzed. The table shows the glycerol biosynthesis and osmotic stress response-related genes
retrieved being differentially expressed across the strains analyzed. The colors used
in the table indicate the percentiles projected on the entire amounts of the transcripts
counted (green stands for 90 %, white for 50 % and red for 10 %). [Note: This table could not be inserted due to technical limitations. It can be found as an image file in the supplemental files section]
The analysis resulted in a set of candidate genes which were found to be upregulated
in both the salt stress samples of the ΔMohog1(adapted) and the wildtype strain, whereas these genes were not regulated in the lof mutant
ΔMohog1 (Tab. 1, yellow marked). Among these candidates, we identified genes encoding the
glycerol H+-symporter MoSlt1p (MGG_09852), one phosphoglycerate mutase (MGG_06642,), one glycerol-3-phosphate
dehydrogenase (MGG_00067 (MoGpd1p)) and one phosphatidyl synthase (MGG_00099 (MoHad1p)).
The HOG pathway is not responsible for adaptation in ΔMohog1(adapted)
We searched for possible interactions between MoHog1p and other osmotic stress responsive
or associative gene products using the SMART website to find links between putative
interaction partners of the MAPK MoHog1p in the adapted strains. The genes identified
were finally used to inspect their expression patterns within our set of DE genes
(STRING analysis, Fig. 9 [A]).
Fig. 9:Interaction network analyzed by the STRING website and gene expression profiles of
HOG1-related genes across the strains analyzed in this study. (A) STRING network visualization showing 30 proteins known to interact with Hog1p
in yeast. With a confidence cutoff of 0.9, the resulting network contains 48 functional
associations between 30 of the proteins. (B) Expression profiles of DE genes obtained
from the STRING analysis across the strains analyzed. (C) Expression profiles of candidate
genes derived from the transcriptome analysis of the strains selected.
As expected, the transcript level of the responsive genes belonging to HOG pathway
were found to be highly upregulated in case of the wildtype strain in response to
osmotic stress (Fig. 9 [B]). Thus, among the genes with the most abundant transcripts
were MGG_01822, MGG_08212 and MGG_08547 encoding HOG1, a BZIP transcription factor and CAMK1 kinase, respectively (Fig. 9 [B], [C]). Meanwhile, none of the HOG1-associated genes, except for MGG_06759 encoding a heat shock protein, were transcriptionally
active in ΔMohog1 and in ΔMohog1(adapted) (Fig. 9 [B]). That leads to the conclusion that other mechanisms operating independently/outside
of the HOG pathway may be responsible for the phenotype observed in the adapted strains.
{kind=link}