Marine sample collection and fungal isolation
Collection of marine sponges were collected from the red sed of Jeddah, KSA. Eight endophytic fungal strains coded as (NAS25, NAS26, NAS30, NAS42, NAS46, NAS49, NAS51, NAS52) have been isolated using potato dextrose media.
Screening of fungi for cellulolytic enzymes activity
The isolated fungal isolates were screened quantitatively for their ability to produces cellulases. Out of eight isolates, 3 isolates displayed positive cellulolytic activity. Fig. 1 displayed both positive
and negative cellulase endophytic fungi on plates flooded with either (a) Iodine solution or (b) Congo red.
the appearance of Clear zones around the fungal colonies indicates
their ability for cellulase production. The obtained result showed that out of 8 fungal isolates only 4 isolates displayed a pronounced cellulase enzymes activity Kasana et al. [26] developed an accurate and obvious method by using Gram's iodine and Congo which gave a more rapid and highly apparent
detection results. Several studies have been used gram iodine for the screening of cellulase-producing fungi [27].
Quantitative screening fungus cellulolytic activity
Based on the qualitative screening results, four endophytic fungal strains have been selected for further cellulase enzymes activity. The assay involved the evaluation of FP-ase, Salicinase, and CMC-ase activity. Results in Table 1 represent the data for the positive selected four fungal strains based on the qualitative screening ‘formation of a clear zone on congo red and iodine plates. The strain NAS51 displayed maximum and highest cellulase activity (FP-ase, β-glucosidase (Salicinase), and CMC-ase enzymes) with activity values (FP-ase, 3.13 U/ml; CMC-ase, 2.52U/ml and β-glucosidase (Salicinase), 0.69U/ml). followed by NAS46, which displayed a pronounced FP-ase activity with 1.09U/ml, while it shows weak CMC-ase activity with 0.58U/ml and approximately no Salicinase activity. The rest two fungal strains showed low cellulase enzymes activity. Several reports have been investigated the cellulase activity of fungi. One of these reports was done by Sri Lakshmi and Narasimha [28], who investigated the cellulase enzymes production by four fungal strains, their reports showed that one isolate belonged to Aspergillus sp. exhibited the highest cellulases activity (FP-ase, 14.16U/ml; CMC-ase, 64U/ml and B-glucosidase, 0.014U/ml).
Table 1 Quantitative screening fungal cellulolytic activity
Isolate code
|
Incubation
period (days)
|
Cultural Initial
pH
|
Cultural Final
pH
|
Total protein (mg/ml)
|
Enzyme activity (U/ml)
|
FP-ase
|
CMC-ase
|
Salicinase
|
NAS26
|
7
|
6.2
|
4.5
|
1.20
|
0.21
|
0.26
|
0.19
|
NAS30
|
7
|
6.2
|
3.2
|
0.95
|
0.125
|
0.35
|
0.16
|
NAS46
|
7
|
6.2
|
3.4
|
1.06
|
2.09
|
0.58
|
0.13
|
NAS51
|
7
|
6.2
|
4.2
|
1.36
|
3.13
|
2.52
|
0.69
|
Identification of cellulolytic fungus
Phenotypic identification
The fungus colonies are fast-growing, Colonies reaching 3-4cm diameter in 7 days at 28°C, on Czapek, yellow to buff with brown margin, zonate, reverse brown. the conidial heads are radiate. The Conidiophore size is 9.0 µm in diameter. The vesicle is globose with a diameter of 26.0 µm. The primary streigmata7.9 X 3.0µm, secondary sterigmata was 5.5 X 2.2 µm. The conidia are spherical with3.0 µm in diameter. The morphological examination of the selected fungus confirmed that the isolated microbe belongs to Aspergillus sp. as shown in Fig. 2.
Genotypic identification
18S ribosomal DNA (rDNA)-based identification of selected fungus NAS51 considers as an accurate tool to confirm and to identify fungi. The 18S rRNA gene was extracted, sequenced, and analyzed by alignment of the obtained sequence with identified sequences deposited in the GeneBank database using BLAST tool to identify the similarity score and to calculate the statistical significance of the matches http://www.blast.ncbi.nlm.nih.gov/Blast. The result established a very close similarity of the obtained sequence with aspergillus sp. with 100% homology and identity. The phylogenetic analysis and the tree were composed using the Maximum Composite Likelihood method (Fig. 3) by the MEGAX program according to Kumar et al. [29]. Based on the analysis of the DNA sequence and the morphological characteristics of the NAS51 isolate, the strain was identified as Aspergillus sp. NAS51 and deposited in GenBank with accession no. MZ665462.1
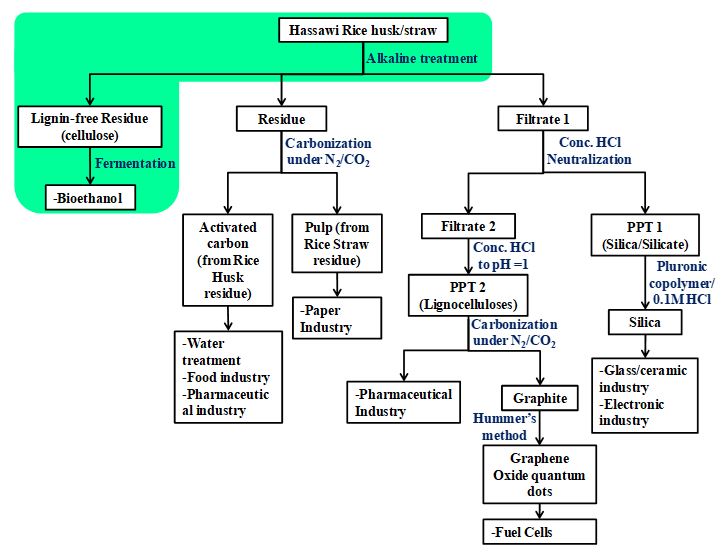
Alkali Pre-treatment of rice straw
Pretreatment with NaOH (1.5%)
To remove the free non-structural sugars before the pre-treatment step, the rice straw was washed with hot deionized water (80o C) then dried in the oven at 45oC. Rice straw was pretreated with NaOH (1.5%) at 121oC and 15 psi pressure for 1 hr., at the ratio of 1:10 to substrate and NaOH solution. The pretreated rice straw was washed with tap water until the pH of the filtrate reached 6. The washed straw was dried at 60oC overnight to constant weight and stored at room temperature for further use (Fig. 4). Table 2 showed the chemical constituents of untreated and chemically pretreated rice straws and weight loss after the alkali pretreatment process.
Table 2 Chemical composition of untreated and alkali-treated rice straw.
Composition
|
Percentage (%)
|
untreated Rice straw
|
Alkali (NaOH)/
autoclaved Rice straw
|
Lignin
|
22.25
|
7.50
|
Hemicellulose
|
26.23
|
15.02
|
Cellulose
|
30.25
|
50.12
|
Ash
|
9.26
|
15.20
|
Loss in weight
|
---
|
25.00
|
Cellulase production by Aspergillus sp. NAS51 via solid-state fermentation and optimization of cultural condition
The strain NAS51 which displayed the highest FP-ase and CMC-ase activity was cultivated on alkali pretreated rice straw and mineral salts as moisture content, the activity was assessed and results showed that at pH 6 and temperature 28 the cellulase enzyme activity of the Aspergillus sp. NAS51 using alkali pretreated rice straw as a substrate under SSF.Extraction of cellulase enzymes was carried out using 0.1 M citrate buffer pH4.8, after the addition of 100 ml of the buffer to each flask, the mixture was put on a rotary shaker for 1 hr at 150rpm. After that, the enzyme was separated from the solid biomass residues by filtration.
Optimization of cellulase production
Effect of pH on cellulase production
The effect of pH on cellulase enzyme production is considered one of the most effective factors that control the fungal strain growth and enzyme production, which controls the transportation of the nutrients through the cell membrane [30]. Several reports have studied the effect of pH of cellulase enzymes produced by fungi. Some fungal cellulases were found to be produced at low pH, while other fungi secrete their cellulases at high pH. Results showed that Aspergillus sp. NAS51producesmaximum cellulase (FP-ase) activity at pH 7.0 with activity (4.50 U/mL)(Figure 5a). the obtained results indicate that, Aspergillus sp. NAS51 produces neutral cellulases. This result is in agreement with a study conducted by Raghuwansh et al. [31] on the mutant strain of Trichoderma asperellum RCK2011 which produces cellulases at a pH range, 4.0–10.0.
Effect of temperature on the production of cellulase enzyme by Aspergillus sp. NAS51
The optimum temperature for the production was measured by incubating the cultural media inoculated with Aspergillus sp. NAS51 at different incubation temp. results in Fig. 5b represent the optimum incubation temperature for maximum cellulase enzyme production. Results revealed that 30oC is the suitable temperature needed to reach the maximum production at pH7 for ten days. The obtained results are in agreement study conducted on Fusarium dimerum and Rhizopus oryzae [32].
Effect of moisture content on cellulase enzyme production by Aspergillus sp. NAS51
Moisture content is an important element for successful enzyme production in the solid-state fermentation process. The increase in moisture level in the SSF process will lead to undesirable results. This fact could be related to the fact that moisture cause swelling of the substrate and makes it easier to be utilized by the organism. low moisture level will be insufficient for microorganisms to solubilize the substrate. While at higher moisture content will lead to a reduction in substrate porosity, and this will limit the availability of oxygen for fungus and consequently will reduce the fungus growth, metabolism, and enzyme production [ref]. Therefore, the influence of moisture content on cellulase enzyme production has been investigated. The results in Fig. (5c) showed that the most suitable initial moisture ratio was70% as greatly enhanced the production cellulase (FPase) to be 4.25 U/ mL. Abdullah et al. [33] found that the optimum moisture content for Aspergillus niger ITBCCL74, when grown in rice straw, was 70%.
Effect of incubation period on cellulase enzyme production by Aspergillus sp. NAS51
Fig. (5d) showed that the maximum cellulase activity was obtained after 11 days of incubation at 30oC, moisture level 70%, pH7. Several reports have evaluated the effect of incubation time on cellulase production under SSF. The maximum cellulase activity by Fusarium dimerum and Rhizopus oryzae was obtained after 9 and 11 days, respectively [32]. While Jafari et al. [34] achieved maximum enzyme activity from A. niger mutant after 10 days.
Effect of nitrogen source on the production of cellulose
Nitrogen elements consider as vital precursors for the construction of protein or enzyme. Fig. (6a) showed the effect of different nitrogen sources in mineral salt medium cellulase (FP-ase) enzyme activity. Results showed that the fungus displayed maximum cellulase activity with 5.62 U/mL when urea was used as a nitrogen source.
Effect of K2HPO4 different concentration
The effect ofK2HPO4 different concentrations was also studied by adding K2HPO4 in a range starting from 1 to 5 g/L. The maximum cellulase activity was achieved at a concentration of 2g/L. while the activity decreased gradually with increasing K2HPO4 concentration (Fig. 6b).
Characterization of crude cellulase enzyme
Optimum pH and pH stability
Cellulase enzyme fromAspergillus sp. NAS51exhibited optimum pH of 7 (Fig. 7a), but it’s very clear that Aspergillus sp. NAS51enzyme can retain about 80% of its activity at pH 8and up to 50% at pH 9. The result obtained showed that the enzyme could work efficiently at a wide range of pH from 6-10. Fig. (7b)showed the enzyme stability after incubation with different Ph at 4oCfor 15 h.
Optimum temperature and temperature stability
This cellulase enzyme showed maximum enzyme activity at 40oC (Fig. 7c), the activity of the enzymes decreased gradually with an increase of incubation. The cellulase enzyme was stable up to 40oC, while it displayed up to 50% activity loss at 50oC, and at 80oC the enzyme retained up to 5% of its activity whereas, it lost its activity over 70oC (Fig. 7d).
Cellulase gene sequencing
PCR amplicon was estimated through agarose gel electrophoresis (prepared with 0.7g agarose dissolved in 1X of 50 ml TAE and 1µl Ethidium bromide 1µg/ml) against DNA ladder and shows a single band opposite to 1900bps (Fig. 8). Using SnapGene software viewer version 4.1.3., this nucleotide sequence was translated into nearly 542 amino acids (Fig. 9) and the predicted protein size of enzyme resulted from amino acid polypeptide was 59.5 KDa. DNA nucleotide sequence of cellulose coding gene was blasted on NCBI using BLASTx for the amino acid sequence that resulted showed that, amino acid polypeptide sequence of our PCR amplicon have a high similarity with nine recorded mRNA polypeptide sequences from the same genus. Cellulase gene sequence showed 100% similarity with gene sequence from the same genus of the tested isolate, this gene was recorded with accession number XM_015545371.1. Kim et al. [35] noted that the expressed protein, CelM2 (novel endo-type cellulose) was purified from the cellular extracts by using Ni-NTA agarose slurry. However, the final polypeptide product of the enzyme was smaller (60kDa) than the predicted molecular weight of 71.5 kDa, while Wang et al. [36]reported that the open reading frame (ORF) for cellulase encoding gene from novel Bacillus subtilis was 1470 nucleotides and encoded a protein of 490 amino acids with a molecular weight of 54kDa
physicochemical properties of cellulases
The physicochemical properties of Aspergillus sp. NAS51was predicted by using ProtParam tool. The physicochemical properties (Table1) show that the molecular weight of the enzyme is (59218.20 Da). The instability index of Aspergillus sp. NAS51 cellulase is (37.96) means that the cellulase enzyme is theoretically stable. The computed Isoelectric point pI value of Aspergillus sp. NAS51 cellulase is (7.16), and the higher aliphatic index of the enzyme (87.94) indicating that Aspergillus sp. NAS51 cellulase is a thermally stable protein. Meanwhile, the negatively grand average of hydropathicity (GRAVY) values for cellulase enzyme indicated its hydrophilicity, (0.134) (Table 3).
Table 3 Summary of the ProtParam data for the Aspergillus sp. NAS51 cellulase models.
Details
|
Cellulase
|
Amino acid residue
|
539
|
Molecular weight
|
59218.20
|
Theoretical pI
|
7.16
|
Positively Charged Residue
|
44
|
Negatively Charged Residue
|
44
|
Total No. Atoms
|
8321
|
Molecular Formula
|
C2658H4145N701O803S14
|
Aliphatic Index (%)
|
87.94
|
Instability Index (%)
|
37.96
|
GRAVY
|
-0.134
|
Modeling the 3D Structures of Enzymes
The amino acid sequences of Aspergillus sp. NAS51 cellulase was subjected to homology modeling via SWISSMODELweb-server to generate the 3D structures of the cellulase enzyme [20](Waterhouse et al., 2018). The cellulase enzyme was constructed using an endo-β-1,4-glucanase from B. licheniformis (35.45% sequence similarity). Fig. 10a illustrates the generated 3D structures of Aspergillus sp. NAS51 cellulase. Furthermore, the I-TASSERweb-server was also used to generate high-quality model predictions of 3D structure (Fig. 10b) and biological function of protein molecules.
Validation of homology model
To evaluate the predicted 3D structure of the homology model, Ramachandran’s plot of the model was constructed to determine the stereochemical quality of the protein structure by analyzing residue-by-residue geometry. The backbone conformation and overall stereochemical quality of cellulase of Aspergillus sp. NAS51 was calculated by analyzing the phi (Φ) and psi (ψ) torsion angles, and the results are illustrated in the Ramachandran plots in Fig. 11.
Determination of binding site
Biological annotations of the target protein were measured by COACH and COFACTOR based on the I-TASSER structure prediction. While COFACTOR uses structure comparison and protein-protein networks to deduce protein functions (ligand-binding sites, EC, and GO), COACH is a meta-server technique that collects various function annotation results (on ligand-binding sites) from the COFACTOR, TM-SITE, and S-SITE programs. According to a prediction by I-TASSER algorithm for the protein 3D structure, 4 ligands(Cellotriose,beta-D-glucose, cellobiose, and Xylopyranose) were predicted as a target for Aspergillus sp. NAS51 cellulase with five binding sites as follow; two binding site for Cellotriose (‘47,58,127,128,173,174,229,271,305,30’ and ‘217,218,253,254,257’), one binding site for beta-D-glucose (‘47,58,305’), one binding site for cellobiose (‘7,127,128,173,227,229,271,305’) and one binding site for Xylopyranose (‘52,58’). This outcome is in agreement with the experimental results of cellulases that preferentially hydrolyze cellulose. Fig. 12 explains the predicted binding sites in complex with ligands.
Enzymatic scarification of alkali pretreatment rice straw and bioethanol production
This step aimed to hydrolyze the alkali pretreated rice straw using cellulase enzyme produced by Aspergillus sp. NAS51 produces a fermentable sugar that can be used for the production of value-added products such as bioethanol. This enzymatic conversion step is required for polysaccharides breakdown into monosaccharides such as glucose, which can be converted into ethanol and other product through a fermentation process. In the present study, Alkali treated rice straw was utilized for cellulase production by aspergillus sp. NAS51. The alkali treatment is a very important step for de-lignification, to facilitate cellulose hydrolysis by cellulase enzymes, furthermore, it permits bioconversion of glucose into ethanol by yeast. Fig. 13a showed the use of Aspergillus sp. NAS51 cellulase for scarification alkali-treated rice straw. The reducing sugars were determined using the DNS method. Fig. 15 showed that at reducing sugar were 16 g/L and glucose: 11 g/L when 5% of treated rice straw was used. Both total reducing sugars, as well as glucose, were found to be increased by increasing the substrate concentration (Fig. 13b). The hydrolysis of 3% w/v alkali-treated rice straw using the fungal culture filtrate of Aspergillus tamari, for 10 hours resulted in yields of 33.56g/l reducing sugars [37]. The enzymatic saccharification of the alkali-treated rice straw was monitored using HPLC analysis. Results in Fig. 14 confirm the presence of glucose in the sample taken after 24 h. Bioethanol production from lignocelluloses wastes such as rice straw is considered as one of the renewable sources for biofuel production. Fig. 17 showed that after glucose fermentation in sugar solution obtained from enzymatic hydrolysis of rice straw by S. cerevisiae, about 6.21% (v/v) (0.454 ml ethanol/g fermentable sugars) Abedinfar et al. [38] have used Mucorindicus and Rhizopus oryzae for fermentation of rice straw and yielded ethanol up to 0.43 g g−1. His results were compared with the corresponding yield by S. cerevisae0.45 g g−1.
{kind=link}