Matrine inhibits the proliferation of cancer in vitro and in vivo
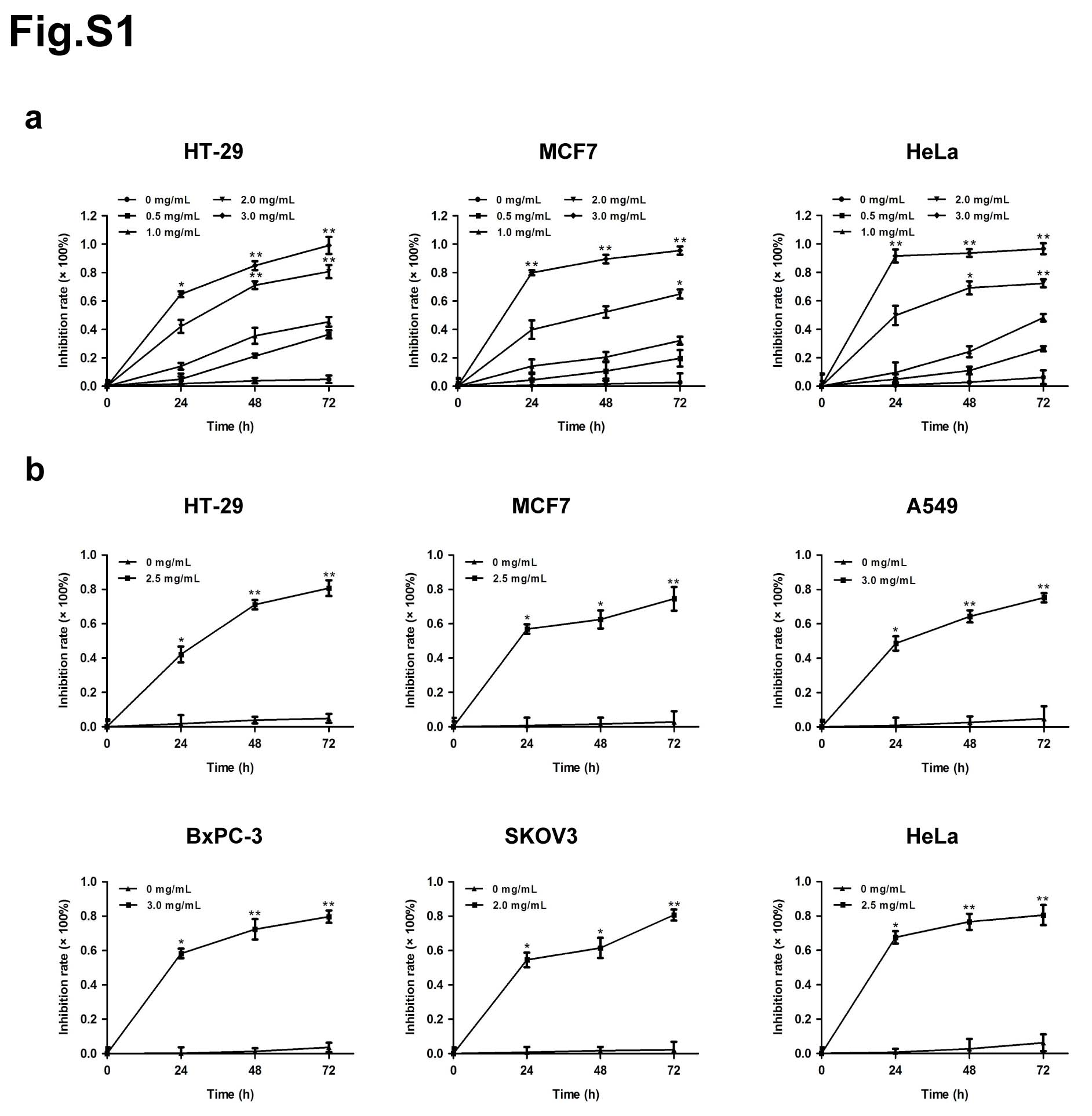
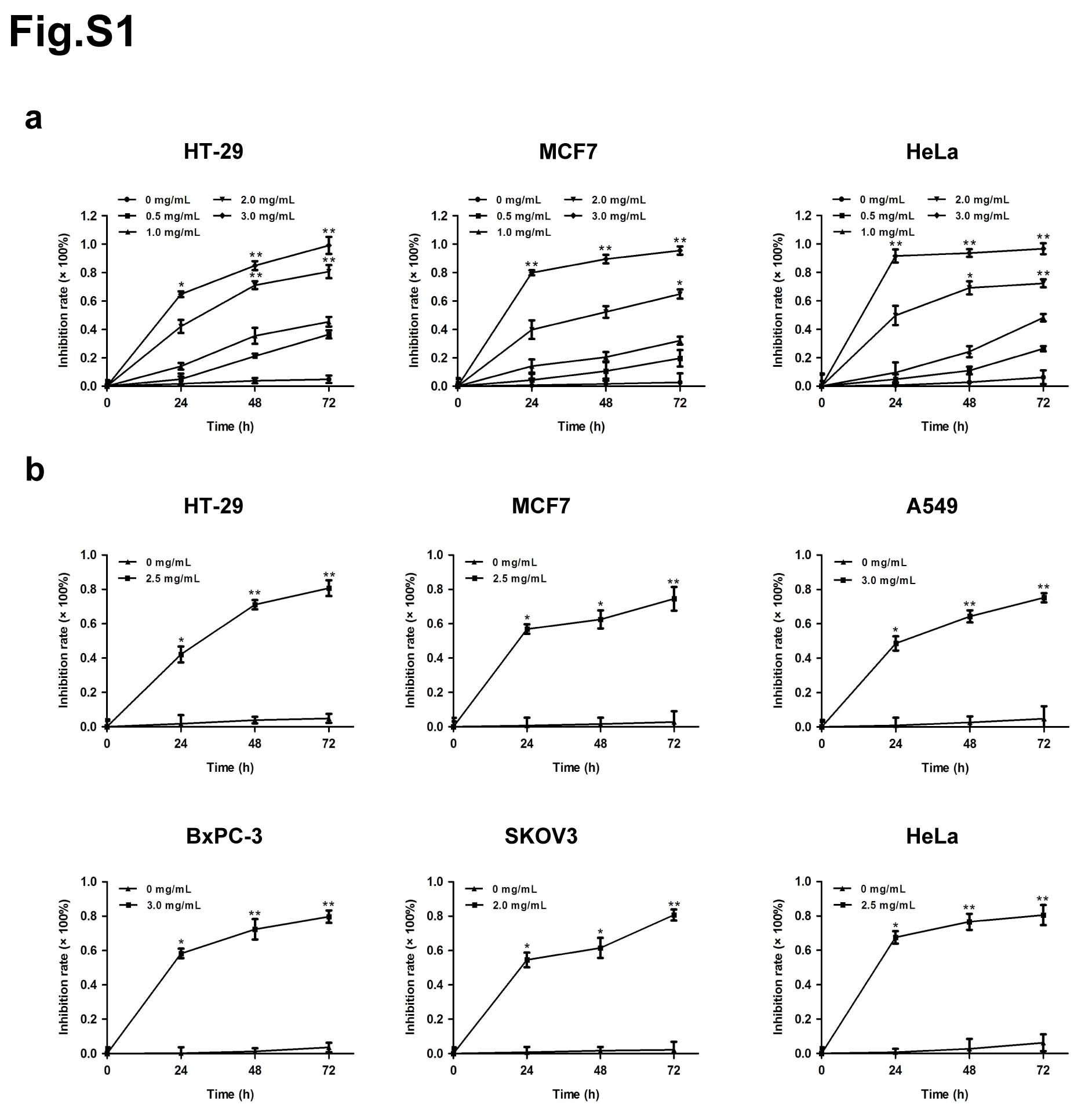
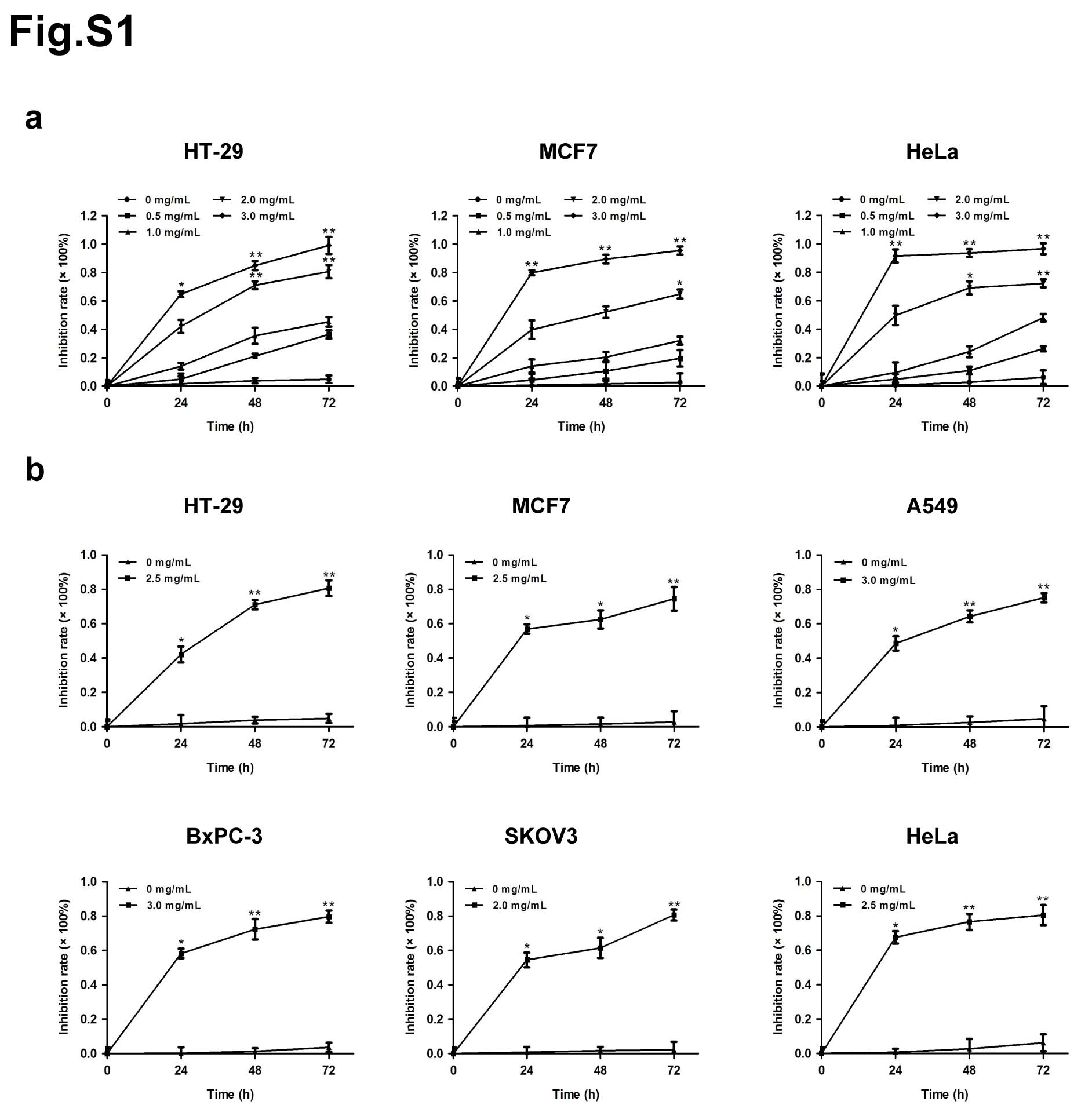
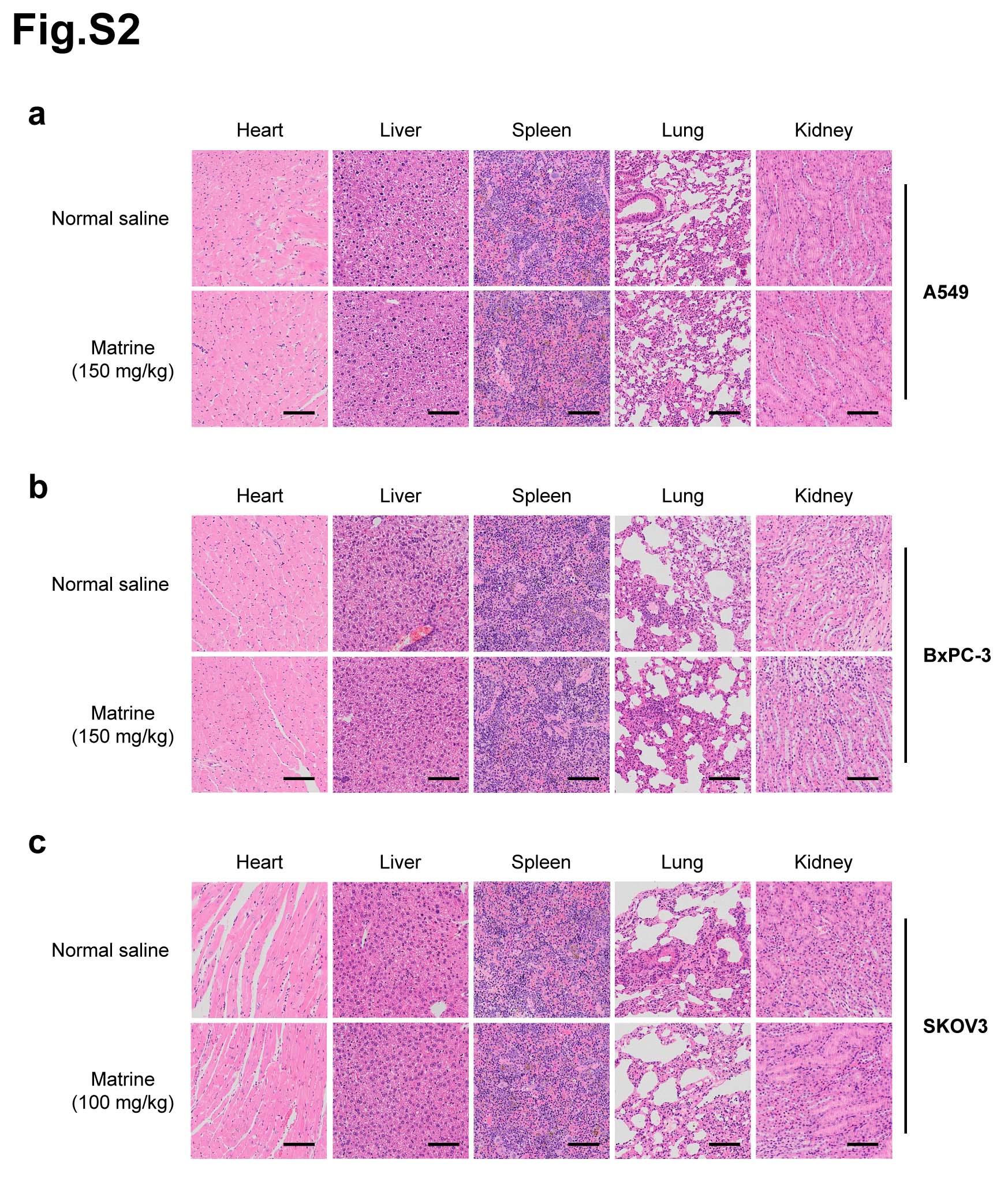
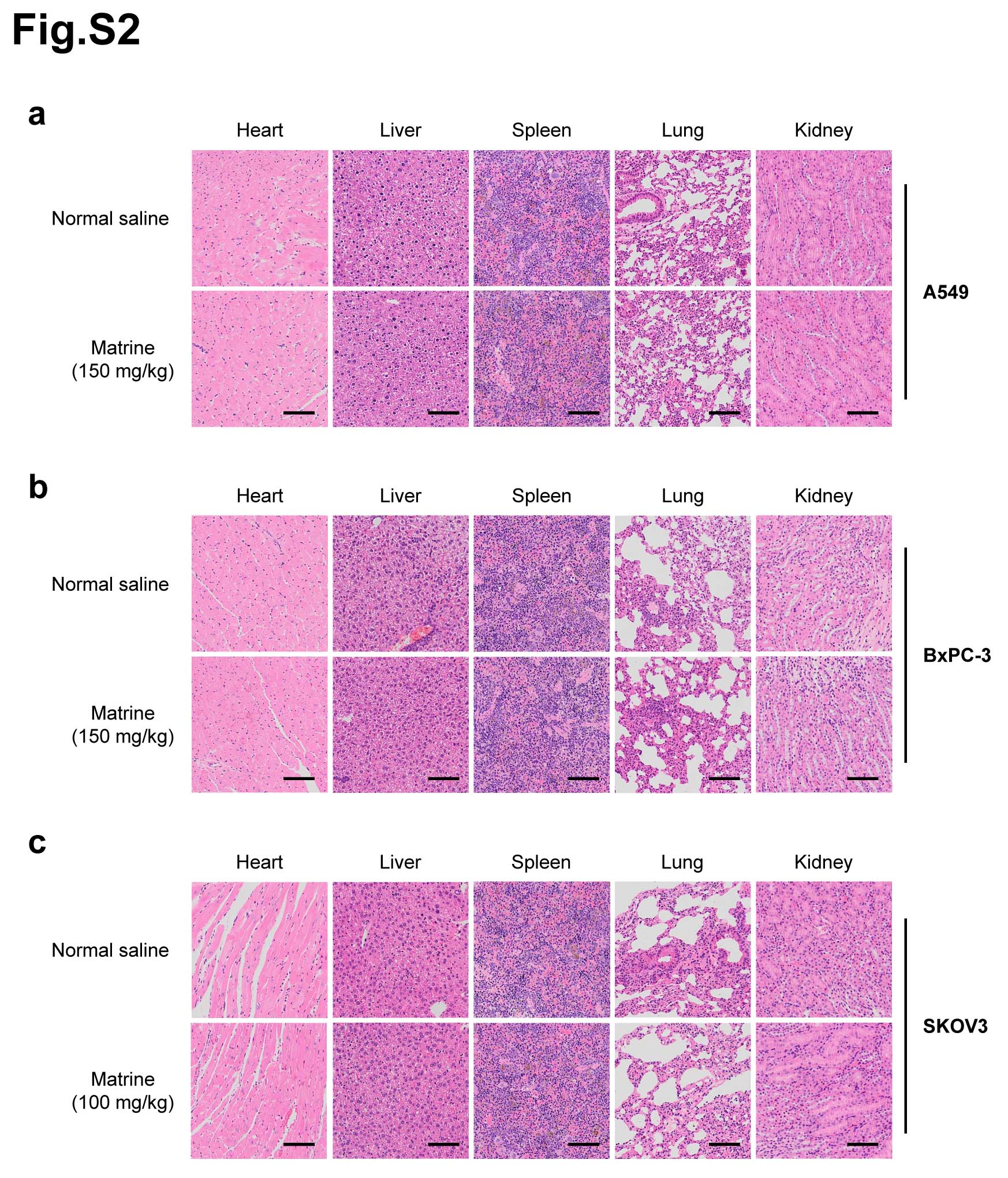
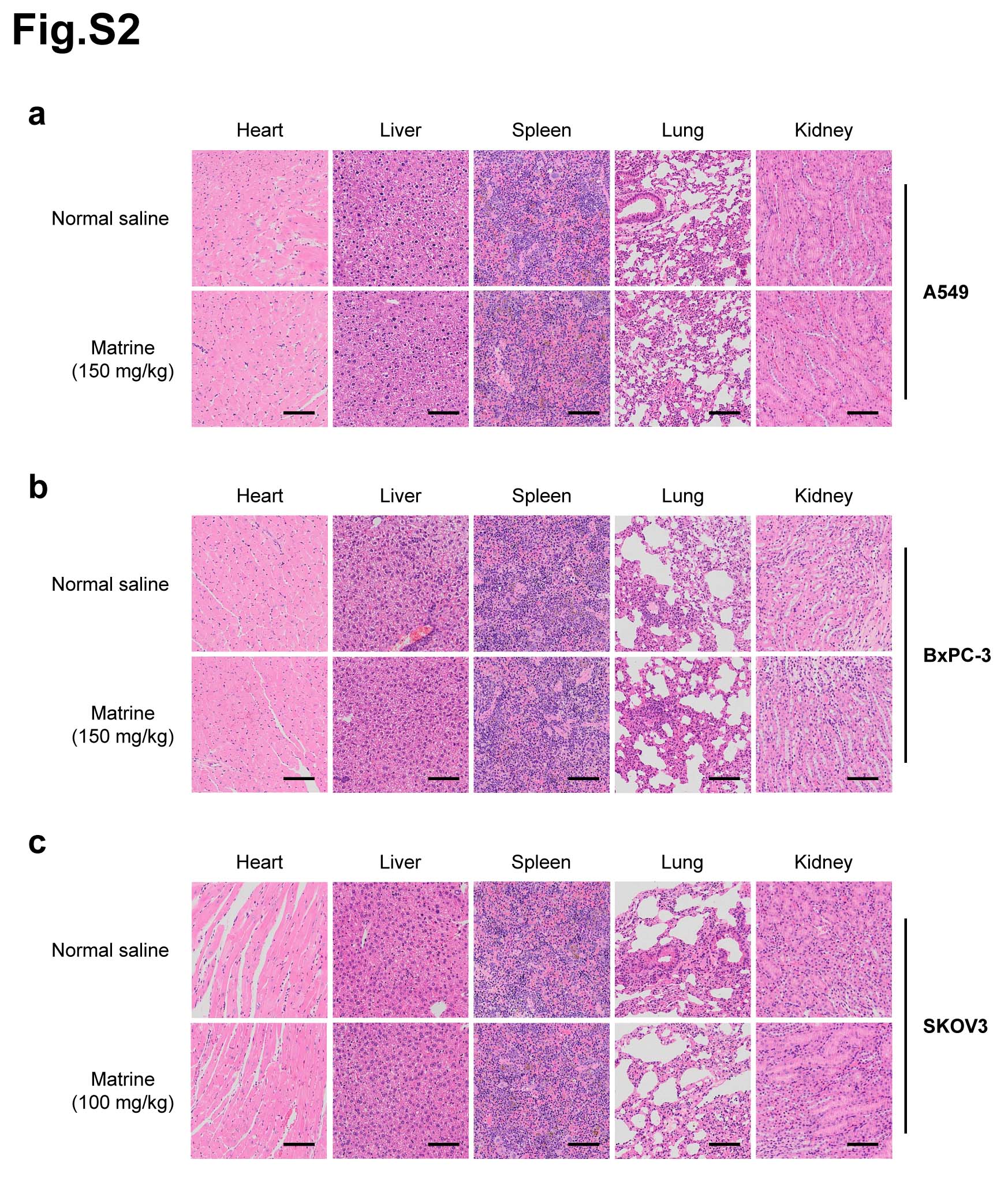
The effects of matrine on the proliferation of cancer cells, including human colon cancer cell HT-29, human breast cancer cell MCF7, human non-small cell lung cancer cell A549, human pancreatic cancer cell BxPC-3, human ovarian cancer cell SKOV3 and human cervical cancer cell HeLa, were determined by MTT assay. Each cell line was treated with matrine at concentration of 0, 0.5, 1.0, 2.0 or 3.0 mg/mL for 0, 24, 48 or 72 h. The results showed that matrine inhibited the proliferation of cancer cells in a dose- and time-dependent manner (Fig. 1a, Supplementary Fig. 1a). The optimal concentration of matrine was 2.5, 2.5, 3.0, 3.0, 2.0 and 2.5 mg/mL in HT-29, MCF7, A549, BxPC-3, SKOV3 and HeLa cells, respectively (Supplementary Fig. 1b). To further determine the anti-cancer effects of matrine in vivo, we injected subcutaneously cancer cells, including A549, BxPC-3 or SKOV3 cells into BALB/c nude mice, respectively. After inoculation, the mice were treated with an intraperitoneal injection of matrine or normal saline thrice a week. The results demonstrated that tumor volume, size and weight of mice in matrine group were found to be much smaller than those in normal saline group (Fig. 1b-d). H&E staining indicated that matrine caused necrosis lesions in tumor tissues of mice inoculated with three cancer cells. IHC analysis showed that treatment with matrine resulted in a remarkably smaller proportion of proliferation marker proteins Ki-67 and PCNA positive cancer cells in tumors compared with the control group (Fig. 1e). Moreover, histopathological analysis of heart, liver, spleen, lung and kidney of matrine-treated mice showed no obviously change compared to the control group (Supplementary Fig. 2), which suggested that matrine could be a safe agent to cancer in vivo. All these data demonstrated that matrine inhibited the proliferation of cancer in vitro and in vivo without obvious toxicity.
Src is the potential target of matrine
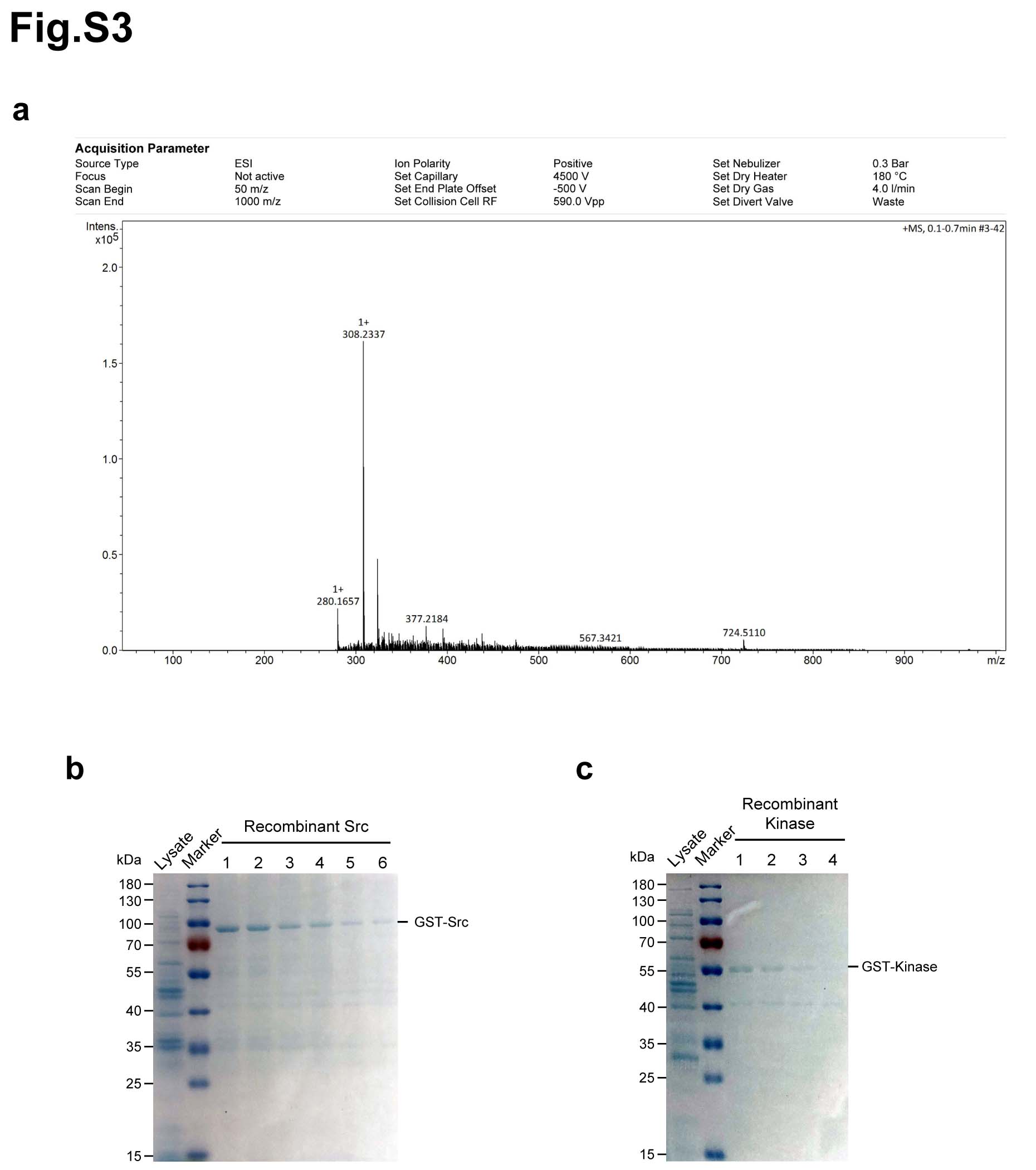
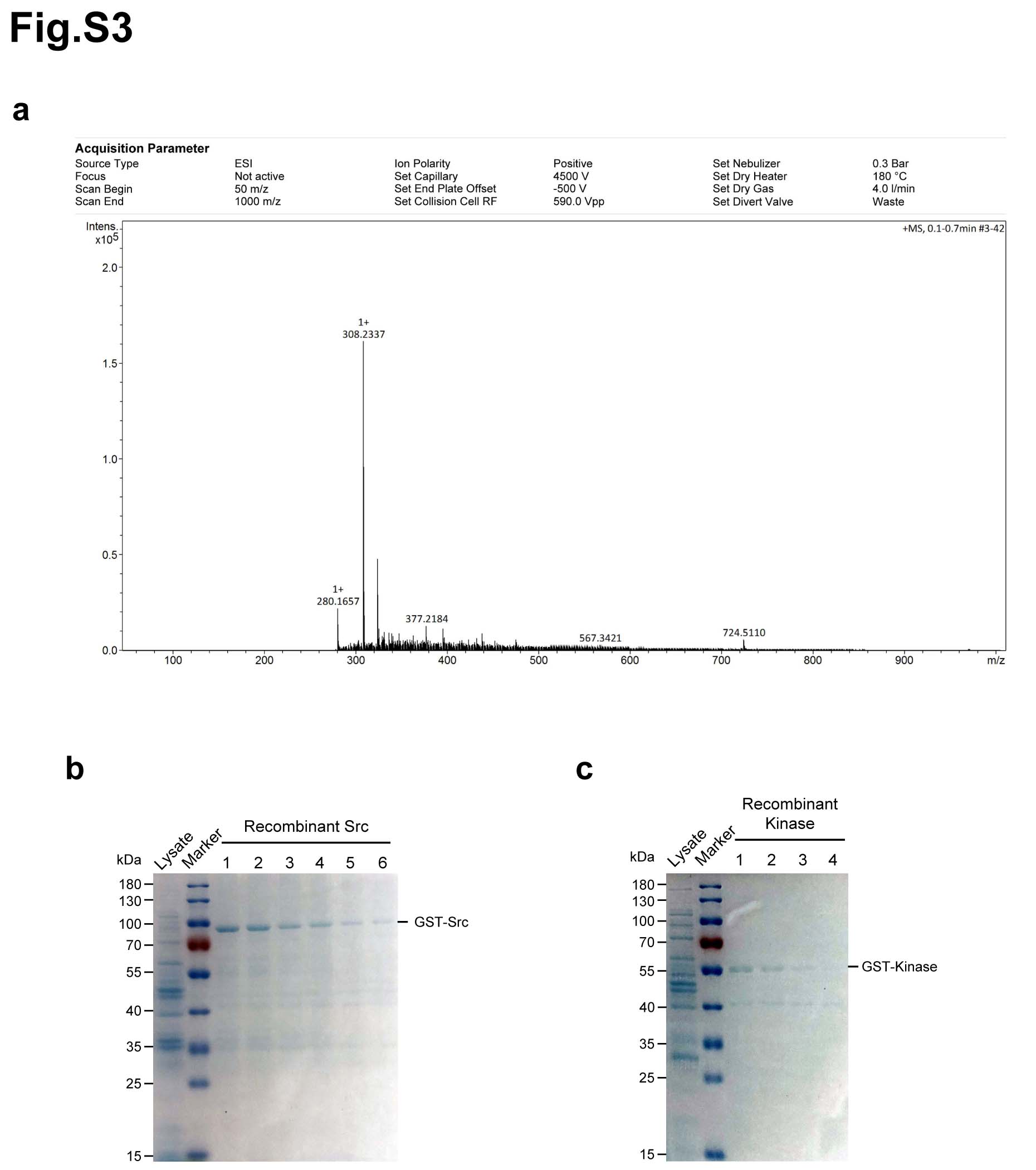
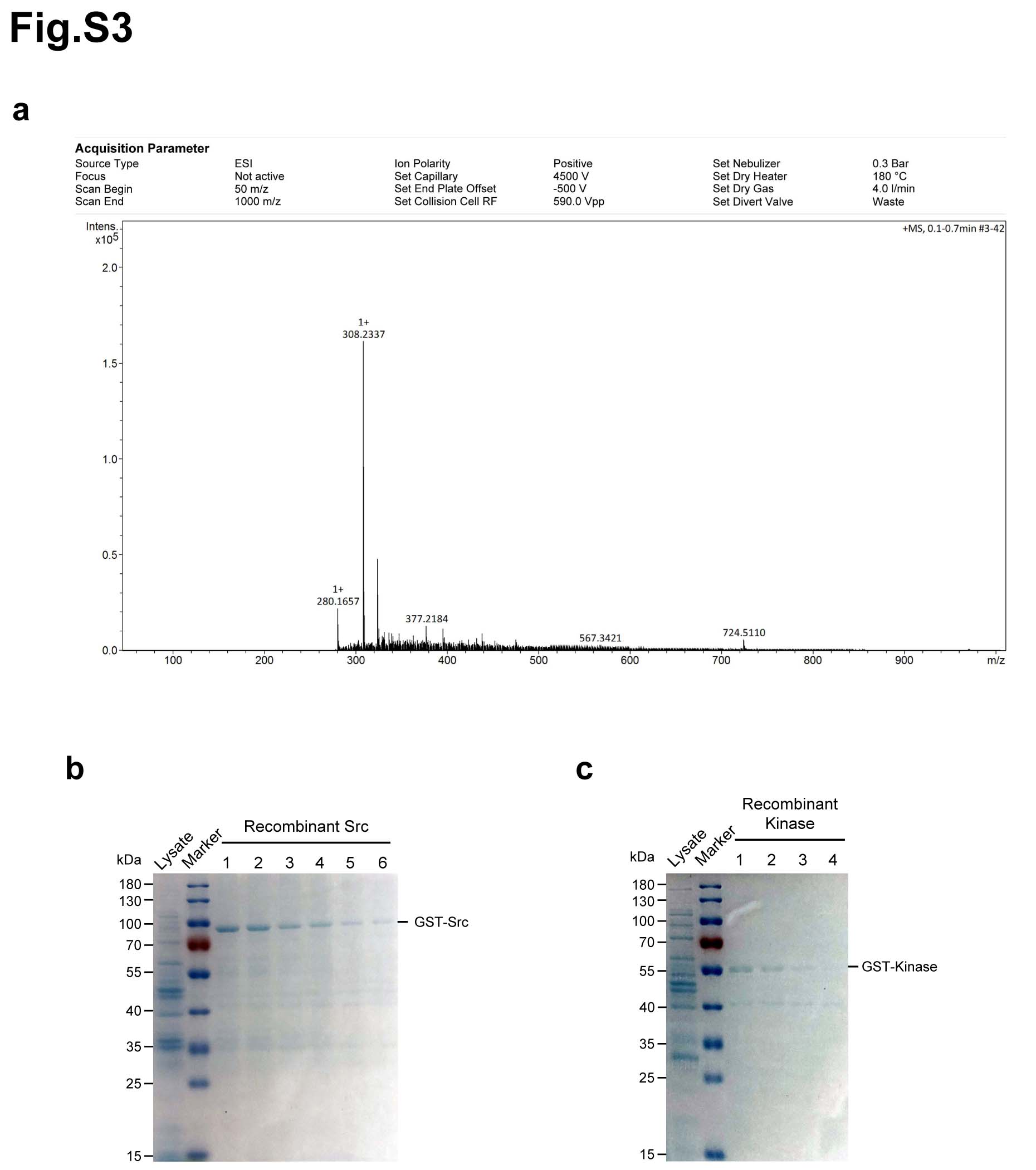
Above data showed that matrine is a potent inhibitor of cancer growth in vitro and in vivo. Then, we sought to explore the potential targets of matrine that mediate its growth inhibitory capability. Matrine and Sophocarpine are alkaloids extracted from the traditional Chinese medicine Sophora flavescens Aiton (Fig. 2a). To explore the potential targets of matrine, we prepared chemical probes for affinity purification. Sophocarpine, the structural analog of matrine, was used to synthetize 13α-(2-amino) ethoxymatrine (P1) (Fig. 2b). High-resolution mass spectrum (HRMS) of P1 was gathered on a Bruker MicroTOF-Q III LC-MS instrument operating in electrospray ionization (ESI) (Supplementary Fig. 3a). As shown in Fig. 2c, matrine-amino coupling resins (MA beads, P3) were made by coupling reaction of 13α-(2-amino) ethoxymatrine with aminolink coupling resins, which were used to pull down the cellular targets of matrine. In addition, competition assay with matrine-amino coupling resins was carried out by adding matrine during coupling reaction. The proteins retained by MA beads were separated by SDS/PAGE and visualized by Coomassie brilliant blue staining. The results showed that one obvious protein band could be observed between 55 and 70 kDa in the group with MA beads but not in group with control beads. Moreover, this protein band was found much weaker with an excess amount of matrine for competition (Fig. 2d). The band in the gel was excised and identified by LC-MS/MS. Mass spectrometric analysis verified this protein as Src protein (Supplementary Table 1).
To further confirm that Src is the direct target protein of matrine, affinity pull-down assay was performed with matrine-amino coupling resins (MA beads) and matrine. As shown in Fig. 2e, Src protein could be precipitated by MA beads, and increasing amount of matrine competitively inhibited the binding between Src and MA beads in cell lysates and tumor tissue lysates. Meanwhile, recombinant protein GST-Src could also be precipitated by MA beads, and competitively inhibited by addition of matrine in immunoblot and sliver staining assays. Furthermore, DARTS results showed that the presence of matrine partially prevents pronase-mediated digestion of Src. Src protein is mostly digested in the absence of matrine, whereas most of them is undigested in the presence of matrine in cell lysates and tumor tissue lysates. GAPDH was used as a negative control, and it was not affected by matrine (Fig. 2f). As ligand may contribute to the stabilization of its target proteins, CETSA was carried out to confirm the interaction between Src and matrine in vitro. The thermal stability of Src protein in cell lysates (Fig. 2g) and tumor tissue lysates (Fig. 2h) was tested by Western blot at the temperature range of 42-64℃ after exposing to matrine or PBS. The results showed that Src was still obviously detectable at the temperature of 60-64℃ in the group exposed to matrine, but not in the control group. Matrine treatment efficiently protected Src protein from temperature-dependent degradation. The significant shift in melting temperatures of Src protein and hence stabilization upon addition of matrine indicated that matrine directly bound to Src protein (Fig. 2i, j). Overall, these results suggested that Src was the direct target of matrine in cancer cells.
Src kinase domain is required for its interaction with matrine
From the N- to C-terminus, Src contains an SH4 domain, a unique domain, an SH3 domain, an SH2 domain, an SH2-kinase linker, a protein-tyrosine kinase domain, and a regulatory tail (Fig. 3a). Src segmented plasmids, including pcDNA3.1-HA-USH3, pcDNA3.1-HA-SH4-UD, pcDNA3.1-HA-SH3, pcDNA3.1-HA-SH2, pcDNA3.1-HA-Kinase were constructed and overexpressed in HEK-293 cells, respectively. The corresponding cell lysates were incubated with MA beads to determine matrine-binding region of Src, which then were dissociated from the beads by denaturation and were probed with anti-HA. Pull-down assay demonstrated that protein band precipitated by MA beads could be only observed in cell lysates transfected with plasmid pcDNA3.1-HA-Kinase. The results indicated that matrine bound directly to the Src kinase domain, and its ability to interact with other regions of Src was negligible (Fig. 3b). To confirm that matrine may definitely bind to the kinase domain of Src, binding competition assay was conducted using pcDNA3.1-HA-Kinase plasmid and GST-Kinase fusion protein. As shown in Fig. 3c, Kinase proteins were pulled down by MA beads, which were reduced by adding an excess amount of MA in cells transfected with plasmid pcDNA3.1-HA-Kinase and in GST-Kinase recombinant protein.
To dissect the critical amino acids involved in the interaction between Src and matrine, the molecular docking of matrine with Src kinase domain was predicted by Discovery Studio 4.5 software. 3D molecular dynamics simulation analysis revealed that matrine was embedded in the cleft of the Src kinase domain. Van der Waals forces were also formed between matrine and the residues of kinase domain, including Met343, Ser347, Ile 372, Asp388, Leu389 and Arg390. Conspicuously, a strong hydrogen-bonding interaction existed between matrine and the residue of Ala392. (Fig. 3d, e). Then, Ala-392 in the kinase domain was mutated to Gly to confirm its involvement in matrine-Src interaction. Plasmid pcDNA3.1-HA-Kinase-A392G was constructed and transfected into HEK-293 cells as well as pcDNA3.1-HA-Kinase. Pull-down assay was performed to determine whether matrine binds to mutated kinase domain of Src in the transfected cells using MA beads. The results showed that matrine bound to wild type HA-Kinase, while significantly reduced binding was observed for mutant Kinase-A392G. Moreover, the binding was also attenuated in cell lysates expressing mutant plasmid by addition of MA (Fig. 3f, g). These results suggested that Ala392 in the kinase domain exhibited an important role in matrine-Src interaction.
Matrine is a non-ATP-competitive inhibitor of Src kinase
The tyrosine kinase Src is activated in a large number of human malignancies and plays significant roles in the development of cancers. The biochemical and structural basis for the basal inhibition of Src protein tyrosine kinase activity was largely elucidated at present [15-17]. To investigate the effect of matrine on Src kinase activity, cells were treated with the optimal concentration of matrine, respectively. The results showed that after treated with matrine, the Src kinase activities in cancer cells, including HT-29, MCF7, A549, BxPC-3, SKOV3 and HeLa, were significantly decreased (Fig. 4a). ATP-competitive inhibitors have been developed and some are used in clinical research, whereas their inhibitory activities must be strong enough to compete with endogenous ATP [18, 19]. Moreover, a number of non-ATP-competitive inhibitors have also been reported for kinases [20]. Molecular docking results indicated that both matrine and ATP bound to Src kinase domain (Fig. 4b, c). To investigate the competitive interaction between matrine and ATP, we measured IC50 values of matrine at various ATP concentrations in SKOV3 cells. The dose-response curves of matrine at ATP concentrations of 25, 50 and 100 μM are indistinguishable. The corresponding IC50 values are 1.83 ± 0.57, 2.15 ± 0.34, 2.05 ± 0.63 mg/mL. This verified that matrine is a non-ATP-competitive inhibitor of Src kinase domain (Fig. 4d).
It has been shown that protein tyrosine kinase could selectively catalyze phosphorylation of the tail tyrosine site in Src kinase domain [21]. Dephosphorylation of Y530, binding of SH2 and/or SH3 ligands, and phosphorylation of Y419 of the activation loop may activate the Src kinase [22, 23]. Cancer cells, including HT-29, MCF7, A549, BxPC-3, SKOV3 and HeLa, were exposed to matrine at its optimal concentrations for 24 h, respectively. The phosphorylation level of Src Y419 was detected by immunoblot. The results indicated that matrine obviously decreased the tyrosine 419 phosphorylation level of Src after matrine treatment in cancer cells (Fig. 4e). Considering that Src Ala392 is adjacent to Y419, we speculated that matrine might block the autophosphorylation of tyrosine 419, resulting in inactivation of Src kinase.
Matrine inhibits phosphorylation signaling pathways in cancer cells
Researches have showed that MEK/ERK signaling pathway plays an important role in regulating cell proliferation, differentiation and apoptosis, thus blocking it can inhibit the proliferation of tumor cells [12]. The expression and phosphorylation levels of MEK1/2 and ERK1/2 in cancer cells were detected by Western blot to evaluate the effect of matrine on MAPK/ERK signaling pathway. The results demonstrated that matrine decreased the phosphorylation levels of ERK1/2 and MEK1/2 in different cancer cells, without change of protein expression (Fig. 5a). The phosphorylation ratios of MEK1/2 and ERK1/2 were significantly reduced in HT-29, MCF7, A549, BxPC-3, SKOV3 and HeLa cells (Fig. 5b). Collectively, the phosphorylation levels of MAPK/ERK signaling pathway in cancer cells were dramatically inhibited by matrine.
The association between the JAK/STAT pathway and tumorigenesis is an important clue of tumor biology. STAT3 is an important transcription factor for gene transcription involved in cell proliferation, differentiation, apoptosis, and angiogenesis in a variety of cells [24]. Sustained activation of STAT3 occurs in many cancers, and can promote tumor growth, survival and progression [25]. To investigate the effect of matrine on JAK2/STAT3 signaling pathway, the optimal concentrations of matrine were applied to treat cancer cells. Immunoblot analysis showed that the phosphorylation levels of JAK2 and STAT3 in cancer cells were down-regulated by matrine (Fig. 5c). The phosphorylation ratios of JAK2 were drastically decreased in HT-29, MCF7 and BxPC-3 cells, and the phosphorylation ratios of JAK2 and STAT3 were obviously reduced in HT-29, MCF7, A549, BxPC-3, SKOV3 and HeLa cells (Fig. 5d). Therefore, matrine could significantly suppress the phosphorylation levels of JAK2/STAT3 signaling pathway in cancer cells.
PI3K/Akt signaling pathway is one of the essential pathways that regulate biological behaviors such as cell proliferation and invasion [26, 27]. Activation of PI3K/Akt pathway is often achieved by its phosphorylation, thus we examined the changes of the phosphorylation levels of PI3K and Akt in cancer cells with matrine treatment. As shown in Fig. 5e, after matrine treatment, the phosphorylation levels of PI3K and Akt in cancer cells were decreased. The phosphorylation ratios of PI3K and Akt were markedly decreased in HT-29, MCF7, A549, BxPC-3, SKOV3 and HeLa cells (Fig. 5f). The results above suggested that matrine significantly inhibited the activation of PI3K/Akt signaling pathway in cancer cells.
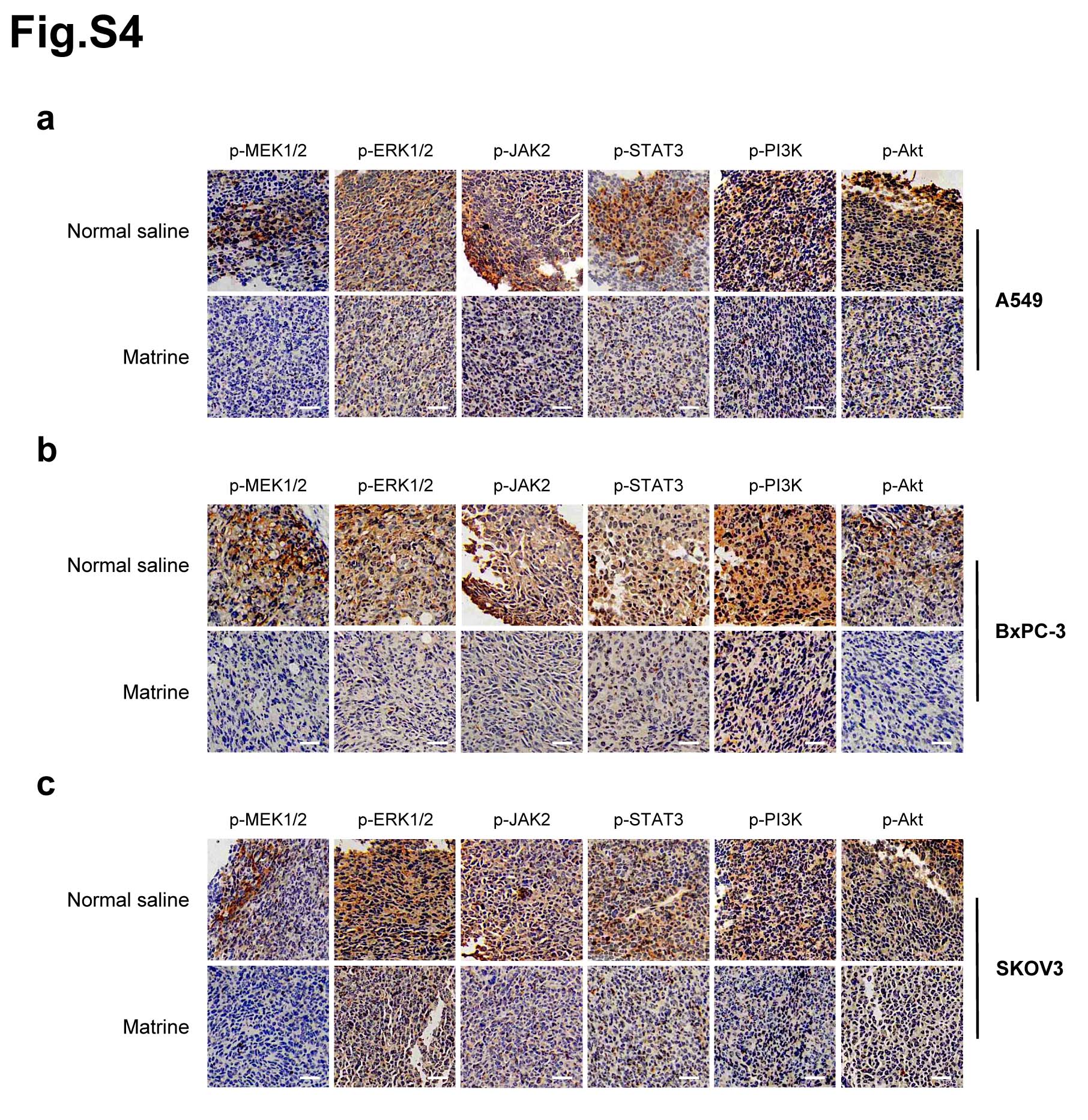
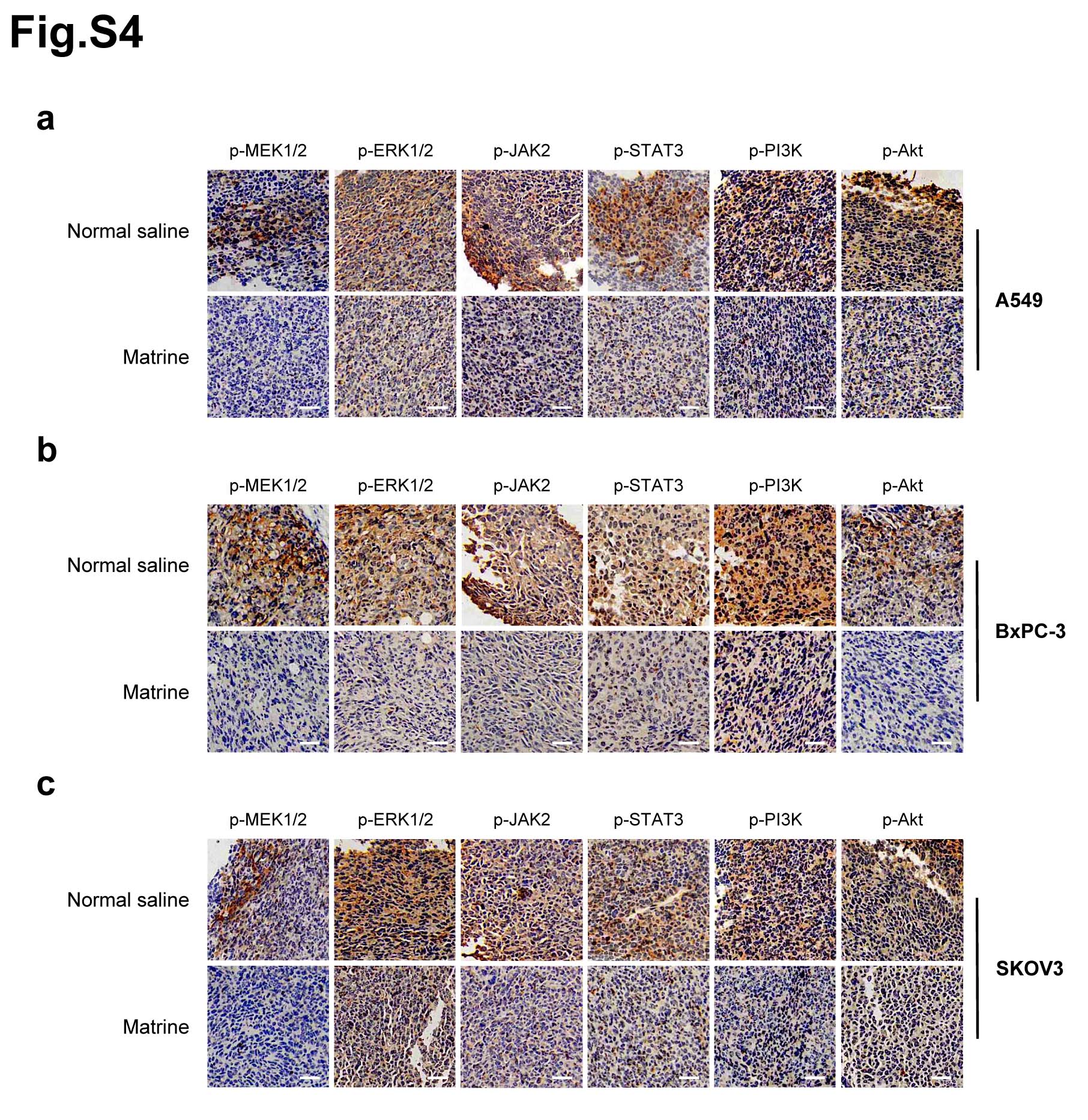
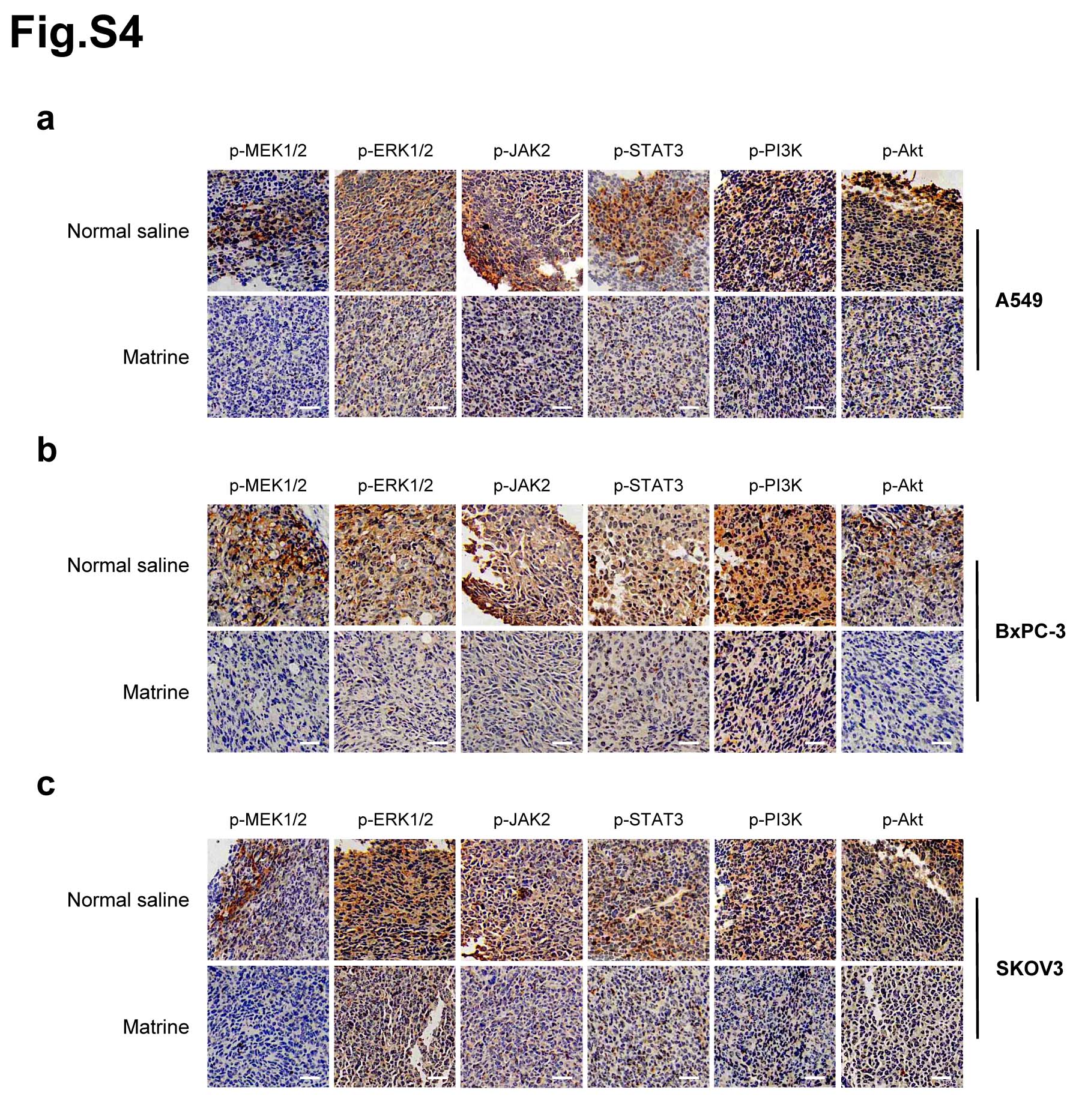
Meanwhile, IHC staining was performed to confirm that the phosphorylation levels of MEK1/2, ERK1/2, JAK2, STAT3, PI3K and Akt in tumor tissues of mice inoculated with A549, BxPC-3 and SKOV3 cells, respectively. The results demonstrated that the phosphorylation levels of these proteins were significantly decreased in tumors with matrine treatment compared with the control group (Supplementary Fig. 4).
Matrine regulates phosphorylation signaling pathways via targeting Src
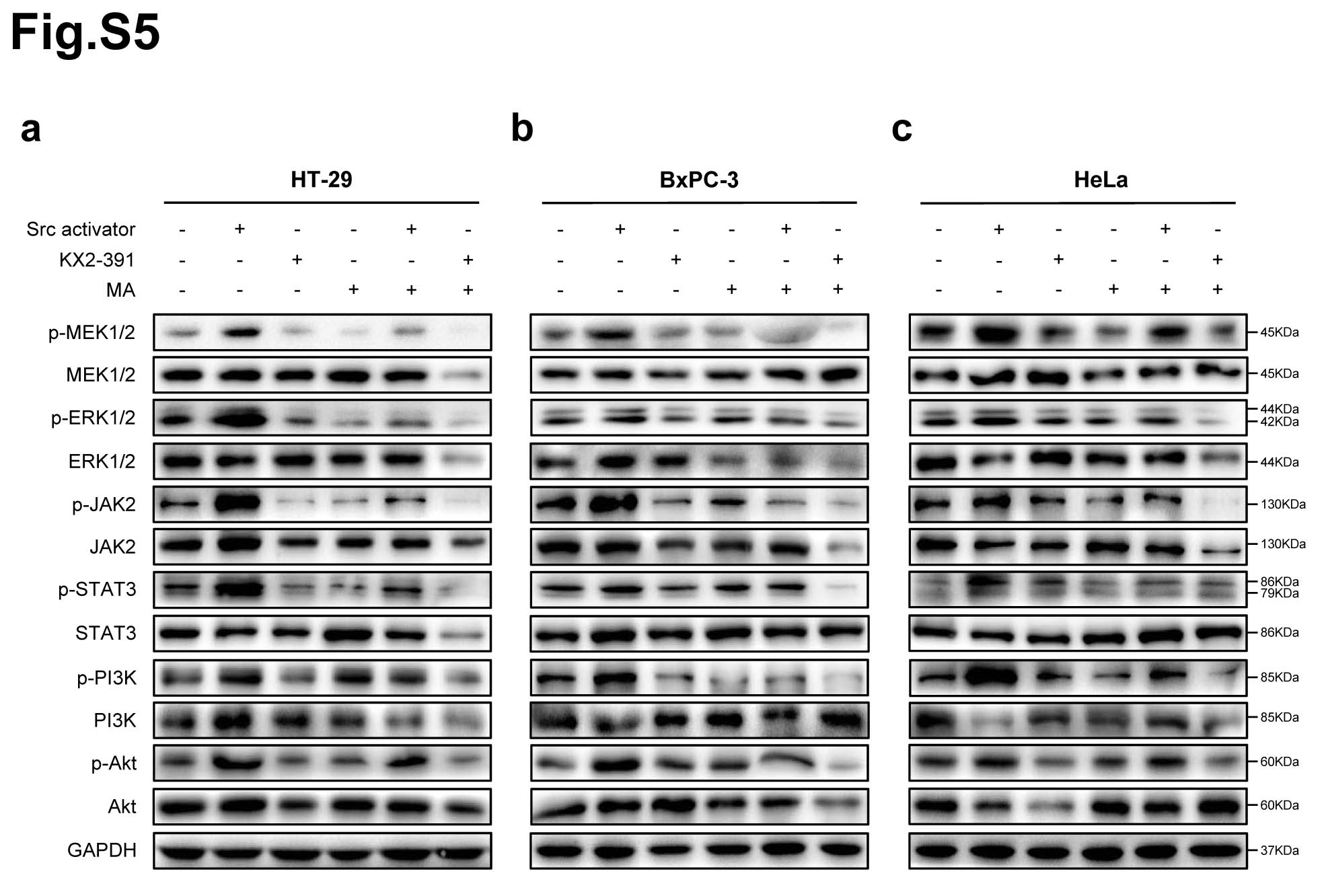
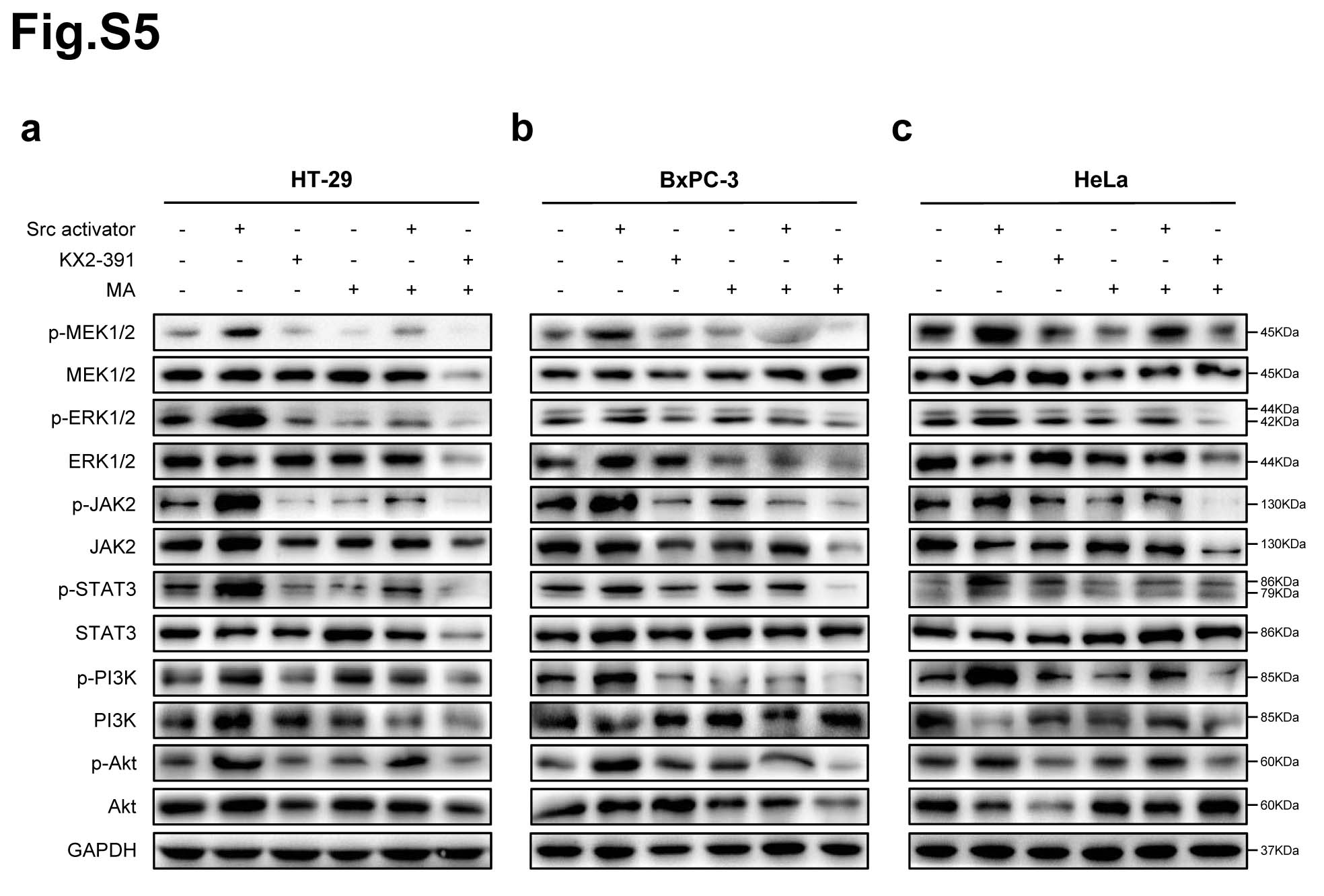
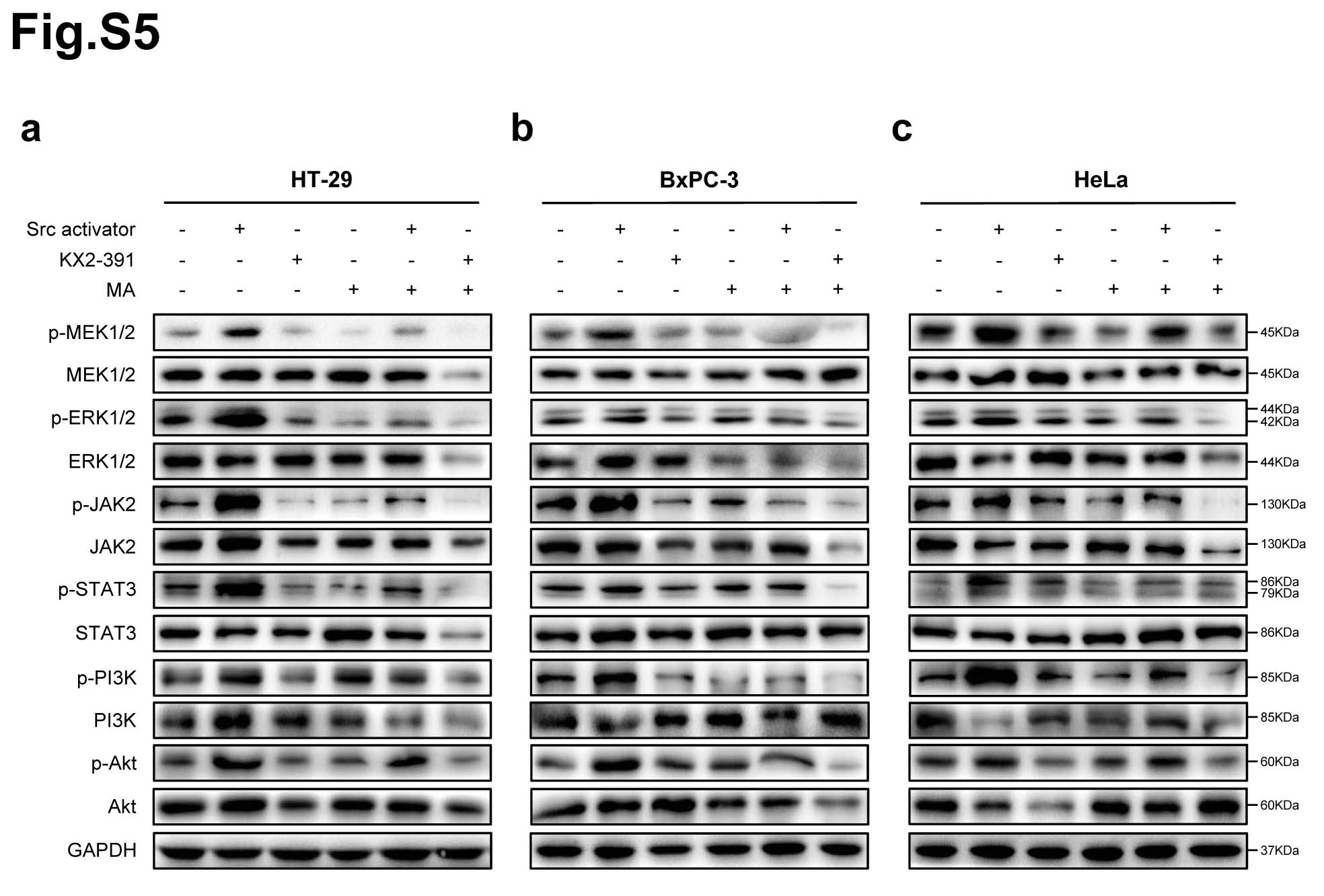
Protein phosphorylation is the most widespread class of post-translational modifications in signal transduction [28]. Src interacts with several protein-tyrosine kinase receptors, such as EGFR, c-Met, PDGFR and IGFR, then participates in pathways regulating cell survival, proliferation and regulation of gene expression [29]. Once activated, Src acts as an upstream regulator of the Ras/MAPK and PI3K/Akt pathways inducing malignant transformation [30, 31]. To further verify that Src is the direct target of matrine in regulating the phosphorylation signaling pathways in cancer cells, Src activator peptide was delivered to cancer cells using a DirectX peptide transfection kit (Panomics). HT-29, MCF7, A549, BxPC-3, SKOV3 and HeLa cells were treated with Src activator or Src inhibitor KX2-391, then exposed to matrine at its optimal concentrations for 24 h, respcetively. The phosphorylation levels of MAPK/ERK, JAK2/STAT3 and PI3K/Akt signaling pathways in cancer cells were examined, respectively. The results demenstrated that Src activator peptide increased and KX2-391 decreased the phosphorylation levels of MEK1/2, ERK1/2, JAK2, STAT3, PI3K and Akt in cancer cells. Their protein phosphorylation levels were further inhibited after exposed to matrine (Fig. 6a-c, Supplementary Fig. 5).
Subsequently, human cervical cancer cell HeLa, gefitinib-resistant human lung cancer cell PC-9-IR (mutant of EGFR, upstream protein of Src kinase) and human pancreatic cancer cell AsPC-1 (mutant of Ras, downstream protein of Src kinase) were transfected with plasmid pcDNA3.1-HA-Src, then treated with matrine or KX2-391, respectively. The phosphorylation levels of MEK1/2, ERK1/2, JAK2, STAT3, PI3K and Akt in cancer cells were detected by Western blot. The results showed that both matrine and KX2-391 could inhibited the phosphorylation levels of these proteins in HeLa cells and EGFR mutant PC-9-IR cells. Consistent with our expectation, Src overexpression may rescue the phosphorylation levels of these proteins, and adding an excess amount of MA may decrease their phosphorylation levels again (Fig. 6d, e). Meanwhile, the protein phosphorylation levels of MEK1/2, ERK1/2, JAK2, STAT3, PI3K and Akt had no significant change by matrine or KX2-391 in Ras mutant AsPC-1 cells, and Src overexpression, matrine, KX2-391 or excess amount of MA had no obvious effect on their phosphorylation levels (Fig. 6f). All these data solidly proved that matrine inhibited the proliferation-related phosphorylation signaling pathways via targeting Src.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}