Although new strategies have significantly improved chemotherapy outcomes in pediatric ALL patients, the prognosis is still poor amongst adults and infants [19]. Identifying a natural compound with minimum toxicity and high efficacy will help develop a novel therapy, which might improve the response to currently available therapies. There are numerous studies regarding the effect of EGCG on cell viability or apoptosis in leukemic cell lines while its effect on autophagy or cross-talk between autophagy and apoptosis in leukemia has not been previously explored. Here, we focused on B lymphoblastic leukemia and studied the effect of EGCG on autophagy, apoptosis, and their interaction in NALM-6 cell line.
The anti-proliferative and cytotoxic effect of EGCG has been investigated in different cancer cell lines, animal models, as well as clinical trials [20]. It has been stated that this compound inhibits proliferation and induces DNA fragmentation in different human leukemic cell lines in a dose-dependent manner [21]. In the present study, we showed that EGCG significantly reduces NALM-6 cell viability (P<0.0001) (Fig. 1).
It is now well established that apoptosis induction is a crucial approach in cancer therapy. Cornwall Cull, et al. have indicated that EGCG results in apoptosis in B-CLL and T-CLL cells in a dose-dependent manner, while it does not affect normal B and T cells [22]. Furthermore, it has been observed that EGCG induces apoptosis in Jurkat cell line through the expression of Fas and increasing Caspase 3 levels [23]. It has been also shown that EGCG prompts apoptosis in leukemic cells such as KG-1, TH-P1, and PML/RARα leukemic mice [24,25]. The results of our study also showed that EGCG treatment results in apoptosis in NALM-6 cell line in a dose-dependent manner, which is in accordance with the study of Fatih M. Uckun, et al. showing that EGCG 10 µg/ml and 50 µg/ml µM caused about 22.5 and 67.8% apoptosis in a 24 h duration in NALM-6 cell line, respectively [26]. We observed that treatment with EGCG 45 µM can significantly increase apoptosis after 24 and 48 hours (P<0.05) (Fig. 2), the concentration which also caused a significant rate of proliferation arrest (Fig. 1).
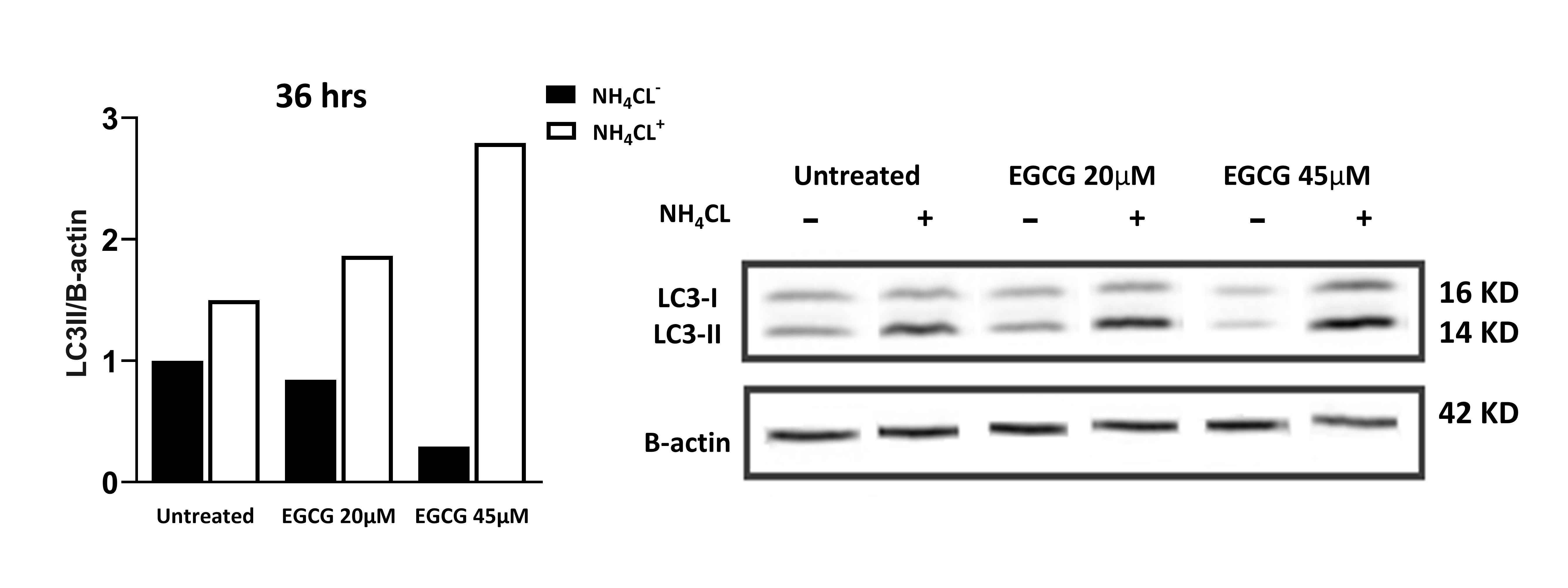
Since autophagy and apoptosis induction pathways are related [27] and autophagy plays a dual role in the pathophysiology of cancer [6], several studies have been conducted to investigate the effect of EGCG treatment on both apoptosis and autophagy mechanisms and their cross-talk. It has been reported that EGCG induces apoptosis and autophagy in five different mesothelium cell lines while treating these cells with EGCG in the presence of chloroquine, an autophagy inhibitor, leads to cell death [28]. Besides, it has been shown that EGCG antagonizes proteasome inhibitors bortezomib toxicity by autophagy induction and protecting PC3 cells from death [29]. However, there is evidence that has demonstrated that activation of autophagic flux by EGCG decreases TRAIL-induced apoptosis in HCT-116, a colon-cancer cell line, by downregulation of death receptors [30]. Interestingly, Jiao Meng, et al. have shown that co-treatment of A549, a non-small cell lung cancer cell line, with EGCG and gefitinib can sensitize them to this drug by autophagy inhibition; hence, this resistance might be due to the induction of cytoprotective autophagy in this cell line [31]. In the present study, we examined the effect of EGCG on autophagy and apoptosis in the presence and absence of NH4CL, an autophagy inhibitor, to investigate how autophagy affects NALM-6 cell survival. Western blot analysis for LC3-II, one of the most important markers of autophagy, showed that EGCG 45 µM significantly induces autophagy in NALM-6 cell line (P<0.05). However, autophagy inhibition by NH4CL didn’t affect apoptosis in the presence or absence of EGCG. In fact, our data are in line with Wong, J et al. that have shown that inhibition of autophagy doesn’t affect apoptosis in NALM-6 cell line, contrary to others who either revealed that inhibition of autophagy increases cell death [15,16] or showing that inhibition of autophagy can be cytoprotective for this cell line [18,14].
Recently, researches have shown that an entire network of transcription factors like FOXO, STAT1, STAT3, NF-κB, and TP53 are involved in the modulation of autophagy. Indeed, several transcription factors including TP53, STAT3, and NF-kB play a dual role in autophagy regulation, through both transcriptional (nuclear interaction) and transcriptional-independent (cytoplasmic interaction) mechanisms, acting as both activators and repressors. In addition, autophagy is regulated by post-transcriptional effectors, including microRNAs, siRNAs, lncRNAs, or post-translational mechanisms such as phosphorylation, ubiquitination, acetylation, as well as histones acetylation [32,33]. Some pieces of evidence have shown that EGCG can affect the expression of downstream autophagy genes by inhibiting Sp1, NF-κB, AP-1, STAT1, STAT3, and FOXO1 or activation of Nrf2 and TP53 transcription factors [34]. It has been also indicated that EGCG participates in gene expression regulation through interfering with the methylation or expression level of miRNAs [35].
In addition, studies have stated that the molecular mechanism of autophagy involves several conserved genes such as MAP1LC3, SQSTM1/P62, Atg2B, and DRAM1 [36]. It has been shown that MAP1LC3 gene expression, with an important role in autophagy induction, may change differently and it is regulated by numerous transcription factors such as ATF4, C/EBPβ, FOXO1, FOXO3, GATA1, and TFEB [37]. SQSTM1/P62 gene also codes an important protein involved in autophagy, transcribed by NF-ƘB, β-catenin, and TFEB [38]. In addition, Atg2B, an essential protein in the autophagy process localizes on the autophagy membrane or the surface of lipid droplets [39]. DRAM1 also known as DNA damage-regulated autophagy modulator 1, transcribed by P53, modulates autophagy and apoptosis [40,41]. It plays an essential role in apoptosis regulation involving BAX and lysosomes [42] and genotoxic stress-induced autophagy [43].
Tumenjin Enkhbat, et al. have shown that treatment of HCT-116 cell line, with EGCG 12.5 µM for 48 h in combination with 2 Gy radiation up-regulated LC3 mRNA. While treatment with EGCG alone caused about a 4-fold change, radiation itself led to an 11-fold change and when they were combined, a change over 15-fold was observed [44]. In contrast, Li, et al. showed that EGCG 20 µM did not alter LC3 mRNA expression in primary hepatocytes, whereas in the pretreated animal models with concanavalin A, a plant lectin that used to induce acute hepatitis, EGCG resulted in a significant decrease in LC3 mRNA level in liver tissue [45]. We also observed that treatment of NALM-6 with EGCG 45 µM for 36 hours led to a significant decrease in the LC3 mRNA level compared to untreated cells (P<0.01), which is in line with the Sainan Li study. Decreasing the LC3 mRNA level may be related to the FOXO1 transcription factor which has been previously shown to be inhibited by EGCG [46].
Regarding P62 expression, Zhong, L. et al. have investigated the effect of EGCG 25 µM and 50 µM on P62 protein and mRNA level in HepG2, a hepatocarcinoma cell line. They observed that EGCG led to the downregulation of the P62 protein level but it did not affect the P62 mRNA level significantly [47]. Li, et al. also have found that EGCG 20 µM did not affect P62 mRNA expression in primary hepatocytes [45]. Nonetheless, it has been reported that the treatment of DIV8 primary rat cortical neurons with EGCG 50 µM for 24 hours caused a significant increase in mRNA levels of P62 compared to DMSO control (P<0.05) [48]. In the present study, we revealed that EGCG 45 µM caused a significant decrease in the P62/SQSTM1 mRNA level (P<0.0005). P62 is one of the Nf-ƘB target genes that can itself be inhibited by EGCG, thus decreasing its mRNA levels [34].
In contrast to LC3B and SQSTM1/P62 genes, there is no evidence concerning the effect of EGCG on Atg2B and DRAM1 genes. We observed that ATG2B mRNA expression level reduces significantly in response to the treatment of NALM-6 with EGCG 45µM after 36 h (P<0.0001). However, it was reported that Atg2B gene expression remained constant in MEFs (mouse embryonic fibroblast cells), in response to starvation after 2, 4, 6, and 8 hours, while, an early upregulation was followed by a later reduction in other autophagy genes and protein markers [49]. Moreover, it has been observed that starvation induces DRAM-1 mediated autophagy in 7702, HepG2, Hep3B, and Huh7 cells. These cell lines were starved for 48 h and DRAM1 mRNA and protein levels had increased significantly [50]. We also observed that treatment of NALM-6 cell line with EGCG 45 µM results in a slight but significant increase in DRAM1 gene expression (P<0.05). Therefore, our data suggest that the reduction of the mRNA levels of autophagy genes and increasing the mRNA levels of DRAM-1 might be related to apoptosis induction with EGCG 45 µM. Nonetheless, observing the decreased expression of autophagy genes, while observing the induction of autophagy upon EGCG treatment in NALM-6 cells, might be due to post-transcriptional or post-translational alterations. Therefore, more investigations are required to find the exact mechanism by which EGCG affects the expression of these genes.
In conclusion, we observed that EGCG decreases the cell viability and induces apoptosis in NALM-6 cell line in a time- and dose-dependent manner. It also results in autophagy induction dose-dependently, and also an alteration in LC3B, P62, Atg2B, and DRAM1 gene expression. Furthermore, inhibition of autophagy does not interfere with the apoptotic cell death induced by EGCG. Considering the importance of autophagy as a secondary cell death mechanism, it would be of great importance to further study the possible involvement of EGCG in inducing autophagic cell death in cells in which the apoptotic pathway is impaired. This compound can also be used in combination with chemotherapeutic drugs to study leukemic cell lines or animal models. Investigating the effect of this combination on autophagy and cell death or proliferation could be beneficial in cancer therapy.
{kind=link}