iBeetle: Large-scale genome-wide phenotypic screen
iBeetle stands for a large-scale systemic RNAi-based screen using the red flour beetle, T. castaneum, as a screening platform. Double-stranded RNA injections were performed in larval or pupal stages and the phenotypic effects scored at multiple levels such as cell biology, physiology, or embryonic and postembryonic development [36, 37]. In the first and second phase of iBeetle, together about 8500 gene models were screened for their function by systematic gene knock-down, which corresponds to slightly more than 50% of the currently annotated genes in this emerged model organism [33]. The obtained phenotypes were documented in iBeetle-Base [38, 39]. The phenotypic analysis included the detection of visible morphological changes affecting the odoriferous stink glands of this beetle, however, without detailed investigation by systematic dissection. Thus, iBeetle serves as a crude first-pass screen to identify potentially interesting genes for a particular biological process. These genes need then to be verified by a detailed re-screening process with a particular focus on the tissue of interest. For 130 genes, corresponding to about 1,5% of the analyzed gene models, phenotypic changes in gland morphology were noted in iBeetle-Base. These were subject to a re-screen procedure based again on RNAi, and for 69 of them (53%), a knock-down gland phenotype could be detected (Additional file 1: Supplementary Table 1). In 60 cases, the original iBeetle-identified phenotype could be confirmed, whereas in nine cases (all from the second iBeetle phase) a distinctive gland phenotype was detected (Table 1). While the iBeetle screen was using a transgenic enhancer trap strain (Pig-19) [41], the re-screen was performed in the wild type San Bernadino strain. The difference in the observed phenotypes might thus be caused by strain-specific differences, which have previously been observed in T. castaneum RNAi-induced phenotypes [42].
Morphological knock-down phenotypes of Tribolium castaneum stink glands
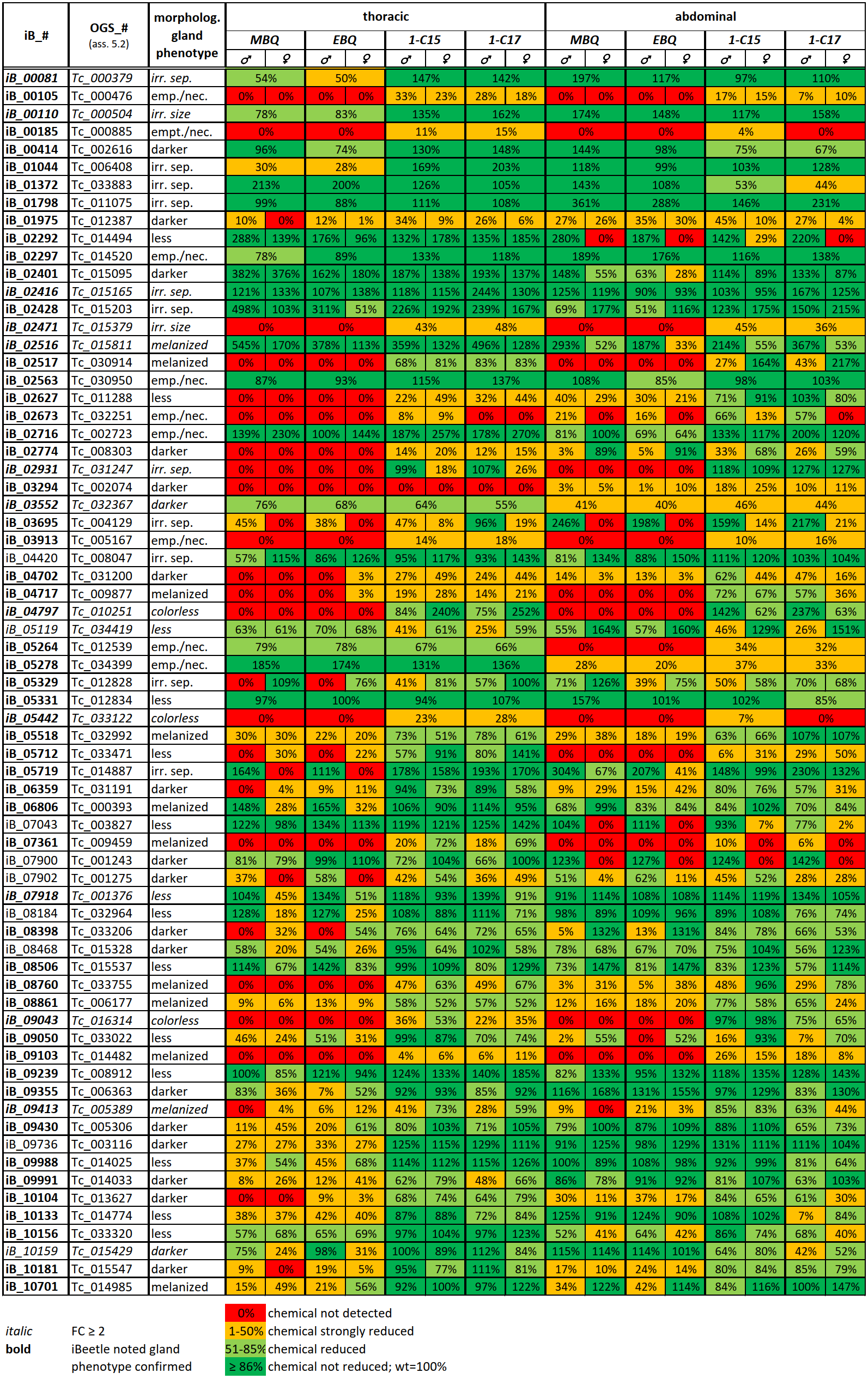
During the first two phases of the iBeetle screen, nine different screeners checked systematically for phenotypes in respect to embryonic and postembryonic development as well as cell biology and physiology. Therefore, the phenotype descriptions noted in iBeetle-Base are quite variable and not completely standardized (Additional file 1: Supplementary Table 1). For the re-screen of all 130 genes with a noted gland phenotype, the visible morphological phenotypes were categorized into seven groups (Figure 1): glands empty and/or necrotic (Figure 1B); reservoir size irregular (increased or decreased, Figure 1E) or containing less secretion (Figure 1H); color of the secretion either darker (Figure 1C), melanized (Figure 1D), colorless (Figure 1F), or showed an irregular separation of the gland compounds (Figure 1G). An additional gland phenotype, that was not observed in the iBeetle screen or re-screen, is turbid secretion (Figure 1I), which was detected in the knock-down of one of the transcriptomics-identified genes (Tc_003768) and originally described as “condensed” (GT12, [40]. The knock-down of this particular gene causes a rare alkene-less phenotype not affecting the benzoquinone production [40], which has so far not been observed for any other gene knock-down. The comparison of the originally described gland phenotypes in the iBeetle screen with the categorized phenotypes of the re-screen is provided in Additional file 1: Supplementary Table 1. For the 69 genes with a re-screen confirmed gland phenotype, the gland morphology category is also indicated in Table 2.
For 61 genes, the iBeetle-Base originally noted phenotype could not be reproduced even with injections of higher concentrations of dsRNA. The majority of the annotated stink gland phenotypes for these genes are hard to identify (‘less secretion’, ‘irregular reservoir size’, and ‘secretion color darker’), as also in wild type beetles, stink gland secretions display natural variation in the degree of filling, color, and shape. Therefore, it is highly likely that these genes were assigned as false-positives in the first-pass iBeetle screen that needs re-screening for confirmation.
Changes in stink gland volatile compounds of gland gene knock-downs
To examine whether the knockdown of the 69 re-screen-identified genes not only caused a visible morphological phenotype but also a change in gland contents, we applied gas chromatography-mass spectrometry to analyze the volatile compounds of the gland secretions independently for the thoracic and abdominal glands. In wild type beetles, the volatile secretion composition is similar in males and females [40], with the four main volatiles corresponding to two para-benzoquinones, 2-Methyl-1,4-benzoquinone (MBQ) and 2-Ethyl-1,4-benzoquinone (EBQ), and two alkenes, 1-Pentadecene (1-C15) and 1-Heptadecene (1-C17). In the different gene knock-downs, the abundances of the four main volatiles were altered to different degrees, reaching from higher than wild type levels, to no alteration, and down to undetectable levels of all or specific compounds (Table 2). The two para-benzoquinones or the two alkenes were usually affected together, whereas the production of the two compound groups seems to be independent. Of the 69 genes analyzed, we found in total that 51 showed strong volatile changes (≥ 50% reduction) in at least one type of compounds (para-benzoquinones or alkenes), in at least one gender or one type of gland. Very strong reductions of both benzoquinones and alkenes in thoracic and abdominal glands independent of the beetle’s sex were observed in the the knock-down of nine genes, which, however, gave rise to very different morphological gland phenotypes (colorless, irregular size, empty/necrotic, darker, melanized). This indicates that the lack of volatile compounds can be the result of very different causes.
Interestingly, 13 gene knock-downs caused specifically a strong reduction of para-benzoquinones levels only, suggesting a specific function in the synthesis of these compounds. In contrast, none of the 69 gene knock-downs led to a specific reduction in alkene production only. In ten cases, reductions of volatile levels were observed to be gland-type specific (six thoracic, four abdominal), and for seven genes the secretion phenotype was found to be sex-specific (two in male, five in female). However, it should be noted that measurements have been done only once with a small number of individuals and gland- or sex-specific effects will have to be confirmed in future experiments. For three genes (iB_04797, iB_05441, iB_09043), whose knock-down caused the morphological phenotype ‘colorless’, also a lack of benzoquinones for both sexes and in both types of glands was observed, linking the yellowish color in wild type stink glands to the presence of para-benzoquinones in the secretions. Besides that, no direct correlation between the visible morphological phenotype and the secretion volatile compound phenotype could be recognized (Table 2).
Encoded molecular functions of genes required for stink gland physiology
To identify the molecular function of the 69 functionally rescreen-confirmed gland genes, the nucleotide or amino acid conserved domains were identified using the respective National Center for Biotechnology Information (NCBI) online search tool [43] and homologs were screened for specifically in D. melanogaster and the entire NCBI nucleotide collection database. The results are provided are provided in Additional file 2: Supplementary Table 2. As expected for a secretory tissue, twenty genes encode enzymes, six transporters, and three channels (Figure 2). However, also two transcription factors were identified as well as thirty diverse other molecular functions, mainly receptors and enzyme regulators. For eight genes (11,6%), no molecular function could be assigned so far, which is probably due to the fact that a systematic genome-wide phenotypic analysis of an odoriferous stink gland tissue has not been performed before.
Transcriptome-based expression levels of iBeetle-identified gland genes
To examine, whether the phenotypically detected genes during the iBeetle screen could have also been identified by a functional genomics approach based on transcriptomics analysis, we identified their expression levels in the published Tribolium stink gland transcriptome [40]. Out of the 130 originally identified genes, only 17 have a more than two times higher expression in the gland tissue (Additional file 1: Supplementary Table 1) and about a third have even a clearly reduced level of expression in the gland tissue compared to a control tissue (Additional file 3: Supplementary Table 3). Thus, the phenotypic screen can also identify genes that are not specifically active in the glands but also in other tissues, but are still necessary for the function of the glands. Of the 69 re-screen-confirmed gland genes, only 14 (20%; Table 1) and of 51 gland genes with a strong secretion phenotype, only nine genes (Table 2) show a two-fold or higher (FC ≥ 2) expression in the gland tissue. This indicates that about 80% of the genes with a function in stink gland physiology have been missed using differential gene expression data to select candidate genes.
Transcriptome-identified gland genes covered by the iBeetle screen
In the odoriferous defensive stink gland transcriptome data, Li et al. [40] identified 77 genes that are highly and specifically expressed in the stink glands (FC ≥ 64). 71 of them were analyzed in gene knock-downs for visible morphological and secretion volatile stink gland phenotypes. Out of these 71 genes, 36 were also analyzed during the first and second phase of the iBeetle screen (Figure 3). Only 29 of the 71 genes showed morphological and secretion volatile phenotypes in the stink glands (Additional file 4: Supplementary Table 4) [40]. Of these 29, 13 were covered in the first and second phase of the iBeetle screen. However, from these 13, only one gene (iB_09413), which caused a melanized phenotype, was confirmedly identified (conf.) also in the iBeetle screen. Two (iB_5763 and iB_5847) were not analyzed (n.a.), since they belonged to the 800 genes that were covered in the pupal screen of the first phase of iBeetle but not in the larval screen, which covered the stink gland analysis. Three genes covered in the first phase of iBeetle caused a lethal phenotype probably due to the larval injection, since the functional analysis of the transcriptome-identified genes was analyzed by pupal injections [40]. For seven genes, the gland phenotypes were not detected (n.d.) in the iBeetle screen despite that fact that they were analyzed. For six of these genes, the phenotypes are hard to detect (less secretion, colorless, turbid, empty/necrotic) and could have easily be missed by the diverse screeners of iBeetle (Additional file 4: Supplementary Table 4). However, one gene (iB_07205) causes a melanized phenotype that should have been detected in the iBeetle screen. In summary, the iBeetle screen identified only one out of eight gland genes that had been identified in the transcriptomics-based approach by Li et al. [40] and were functionally analyzed during the iBeetle screen (Figure 3).
{kind=link}
{kind=link}