Sequence comparison and phylogeny of bugs
The average size fragment of the gene from the amplified bugs was 333 bp. The amplified gene sequences varied from 360 to 388 bp. The consensus gene sequences were blasted in the NCBI Genbank database, and they showed a high degree of homology with the 16S ribosomal RNA gene of C. hemipterus in GenBank at 88.38–90.33% identity. The nucleotide sequences of the 16S rRNA gene from the C. hemipterus are available in the GenBank databases (http://www.ncbi.nlm.nih.gov/genbank/) under the accession numbers MT520974- MT520978. After few processes of removal and alignment of sequences using the T- coffee molecular software, 323 bp of the partial C. hemipterus of the 26 sequences of Cimex sp. and one outgroup, P. humanus were successfully obtained. The overall frequency distributions of nucleotides in the first, second, and third codon positions are as follows: T = 40.7%, 40.3%, and 43.2%; C = 7.5%, 13.2%, and 10.3%; A = 37.5%, 28.9%, and 30%; G = 14.3%, 17.6%, and 16.5%, respectively. Analysis of the sequences revealed a mean GC content of 26.5%. The analysis of the homogeneity of the substitution patterns between sequences using the Disparity Index Test (Monte Carlo Test) with 500 replicates reveals no significant differences across the sequences (Table 1). For estimation of evolutionary divergence between sequences, pairwise distances were computed, and the results are shown in Table 2.
The sequence analysis detected 28 polymorphic sites (16 singletons and 14 parsimony informative sites) among sequences. Results of DNA polymorphism analysis of the five isolates sequences showed that the nucleotide diversity (ð) was 0.57. The total number of mutations (Eta) was 30 while the number of haplotypes (h) was five. The haplotype diversity (Hd) was 1, with a standard deviation of 0.02. The average number of nucleotide differences per site between two sequences (Pi) and G + C content at non-coding positions was 0.04 and 0.26, respectively. Based on the Neutrality test results non-significant Tajimo,s D and Fu,s Fs values were obtained. Genetic differentiation among the population was 0.6. Basic parameters of genetic diversity and neutrality tests are shown in Table 3. Three genomic conserved regions were detected along the 323-bp region (Table 4).
In the phylogenetic tree, the 27 sequences were classified into four large clades (Fig. 1), and the phylogenetic analysis showed that Iranian bugs are grouped with the C. hemipterus. The constructed phylogenetic tree could clearly separate two major clades of C. hemipterus and C. lectularius, although they belonged to the same genus. Interestingly, the tree showed that Iranian specimens were clearly grouped closely together in one branch with a 95% bootstrap support value (Fig. 1). The obtained pattern confirms the validity of 16S rRNA analysis to elucidate the minor differences among closely related species.
Morphology of bed bugs (C. hemipterus)
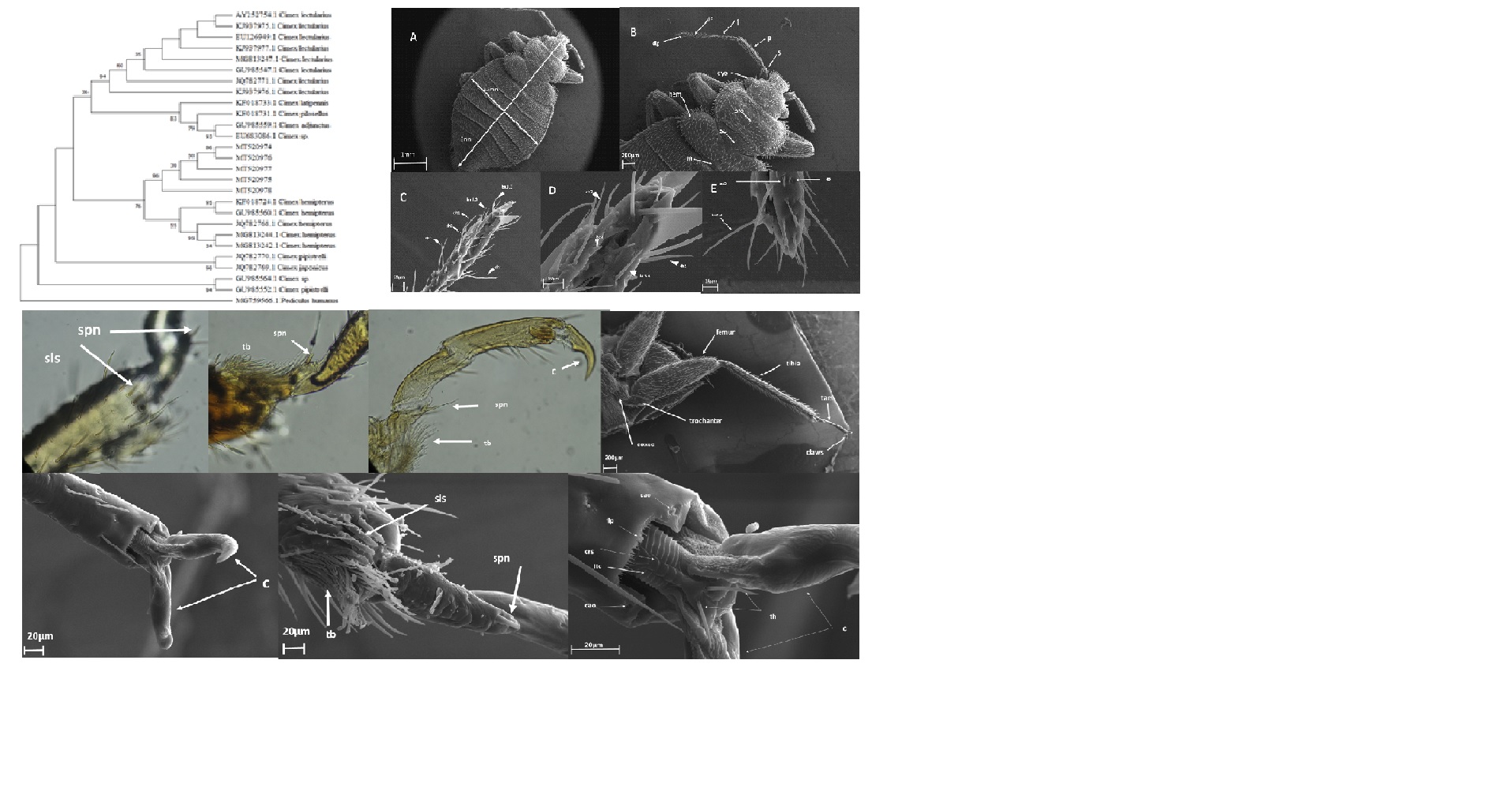
In this study, adult bed bugs were oval and mahogany-colored, and they ranged in length from about 4.6 to 5 mm, with abdomens 2.6 to 2.8 mm wide. The heads were small and cylindrical, with two knoblike multifaceted eyes. Ocelli were absent but compound eyes were well developed (Fig. 2). As Fig. 2 shows, the antennae were four segmented (scape, pedicel, flagellum, and distal flagellum) and placed between the clypeus and eye. The shortest segment was the first segment (scape), and the third and fourth segments were more transparent, slender, and flat than the first and second segments. From the middle of the fourth segment, a deep groove was seen (Fig. 2).
By carefully examining sensilla external shapes and dimensions on the antenna, we could distinguish two types of chaetic, one type of basiconic and one type of coeloconic sensilla in females and males.
Sensilla chaetica type 1 (ch1) were very long, sickle-shaped bristles with longitudinal grooves and blunt tips. The base of the bristles rests on the distal edge of a wide flexible socket and projected from the antennal surface at an angle of 19–96◦ (mean value of 57.5◦) (Fig. 2). The ch1 had the longest pegs, with a mean length of 97.1 µm. Sensilla chaetica type 2 (ch2) were more or less straight bristles, 65–96 µm long, with a hair shaft that strongly tapers from about half of its length and ends in a sharp tip (Fig. 2). The tip was often bent away from the antennal surface. The base of the hair shaft was inserted at an angle of 47◦± 6.5◦ (mean ± std.dev.) into a very tight socket and rested on its concave surface. The surface structure was similar to that of ch1 (i.e., longitudinal grooves present). Chaetica sensilla were the most prominent and longest sensilla on the antennal surface.
Sensilla basiconica (ba) were primarily slightly curved, cone-like hairs, smooth-walled without longitudinal grooves with numerous pores on the cuticular surface, and ended to the blunted tips. The hair shaft inserted basally at an inclination angle that ranged from 19◦ to 40◦ into a socket with a diameter of 2.9–6.4 µm. At its base, the hair shaft had a diameter that ranged from 2.2 to 3.4 µm. Sensilla basiconica were found mainly on both the flagellar segments (Fig. 2).
Sensilla coeloconica (co) were peg-in-pit sensilla, consisting of a small peg set on the floor of a chamber sunken into the cuticle. A small central aperture with a diameter of 2.9–6.5 µm connected the peg inside the chamber with the surrounding air (Fig. 2). Based on the distribution of the various types of sensilla, no sexual dimorphism could be noticed in C. hemipterus.
Mouthparts formed a pointed beak or rostrum that folded under the head in a ventral groove up to the first pair of legs. The mouthparts were modified for piercing the skin and sucking the blood. They consisted of a labrum, a pair of mandibles, and a pair of maxillae. The labrum was short, and it covered the mid-dorsal groove of the rostrum. The labium formed an elongated incomplete hollow tube or sheath called the rostrum. The labium was three-segmented and consists of two lateral lobes and a middle lobe (apical plate) situated on the ventral side. There were sensory structures on the tips of the lateral lobes. The labium was mid dorsally grooved to enclose the four needle-shaped stylets, two mandibles, and two maxillae (Fig. 3). The maxillary and labial palps were absent. Different studies showed that there are a number of differently shaped and differently sized chemo- (gustatory, contact chemoreceptors) and mechanosensilla on the tip of the labium of true bug. In this paper, we are not discussing these sensilla anymore.
The thorax consisted of a narrow canoe-shaped pronotum, a mesonotum (scutellum) which was covered dorsolaterally by reduced fore wings called hemelytral pads, and a mesonotum (scutellum) hidden below the latter (Fig. 3). The prothorax was broad, receiving the head in its concave anterior margin, with the sides extending wing-like laterally (Fig. 4). The same as what Usinger (1966) explained about C. lectularius we found that in the ventral side of C. hemipterus the widely separated sternal apophyseal pits lied between and were concealed by the coxae [19]. The sclerotized area between the pits and partially surrounding them was certainly sternal. The trochantin appeared as a small, detached, sclerite in the membrane anterior to the coxae. The mesothoracic spiracle lied in a small plate just posterior to the coxae. On the ventral side, the mesothorax was almost completely sclerotized, only slight areas about the coxal foramina remaining membranous. At the margin of the body a narrow, sclerotized isthmus connected the sclerotization of the mesothorax and metathorax. In this isthmus, the spiracles of the metathorax lied close to the coxal condyle of the mesothorax. Widely separated between the coxae were the sternal apophyseal pits, which were borne within narrow sclerotized arms branching from the median sclerotized area.
The metathorax was in general similar to the mesothorax in the arrangement of its ventral sclerotization, but the encroachment of the coxae of the mesothoracic legs has narrowed its lateral positions.
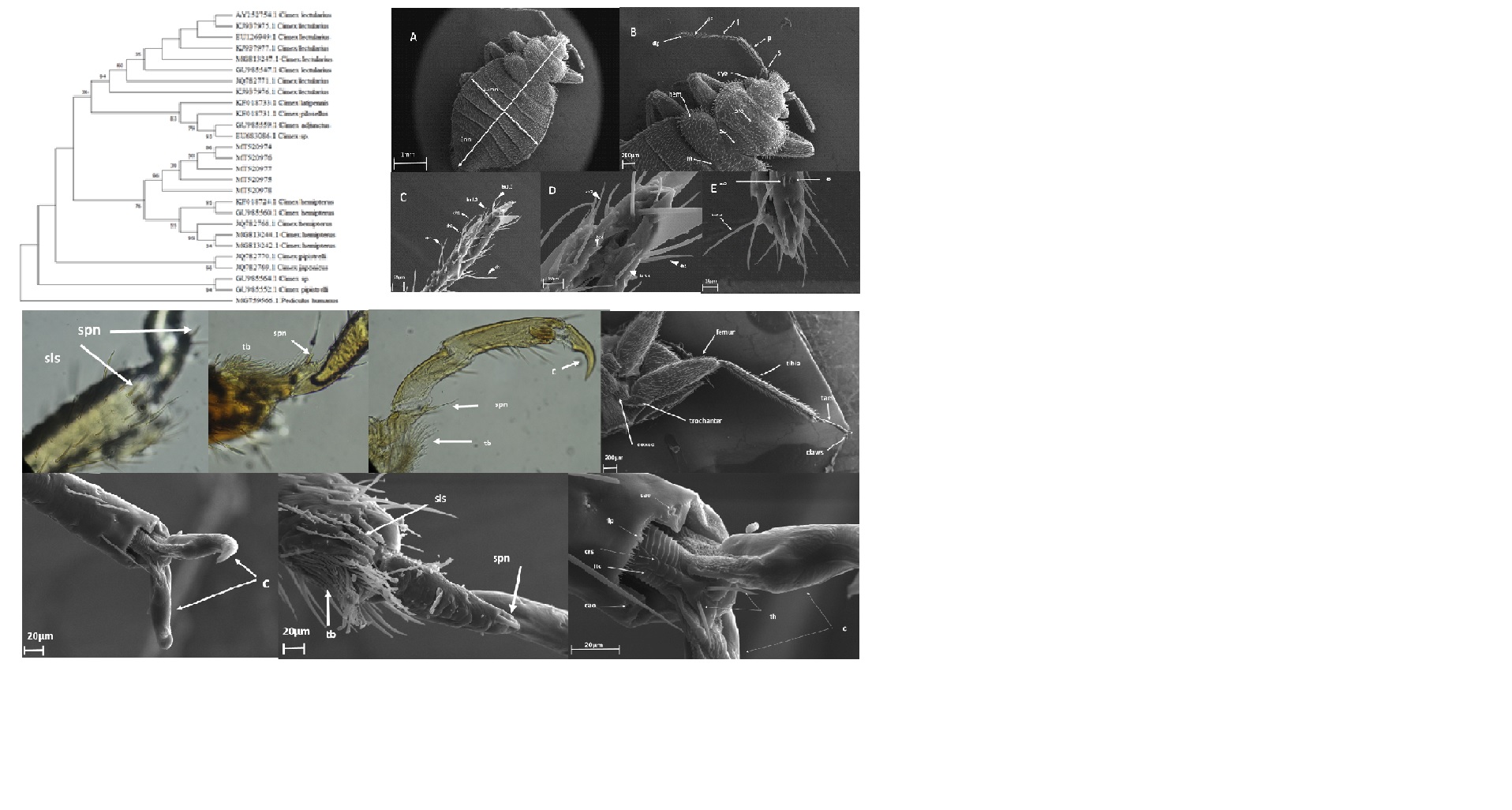
All the parts of the legs were always present except pulvilli and arolia. On the ventral surface of each leg, a tuft of setae was seen on a pad-like extension at the apex of the tibia. The tibia brush varied in length from 35 µm to 60 µm. The pad-like extension at the apex of the tibia had a very wrinkled surface which may be related to glandular openings. In the present study, we found that trichoid sensilla were disseminated among the tibial brush setae.
There were three tarsal subsegments (or tarsomeres) and paired pretarsal claws on each of the six legs. The terminal segment was about as long as the other 2 together. Little is known about the ultrastructure of C. hemipterus legs. Figure 4 shows several structures in pretarsus and as seen in Fig. 4 open-closed system was found in C. hemipterus legs. The open-closed system was formed by the central rack system (crs), two lateral telescopic columns (ltc), and finger-like processes with possible sensory function (flp) (Fig. 4). The rack system could be a system to lock the position of the claw, as was the pad-like structure and always two spiniform setae were in its base.
The abdomen was 10 segmented. The 1st and 2nd abdominal segments were fused. In adult male bug, the abdomen was narrower, and its tip was curved and slightly more pointed than in the female. The intersegmental membranes were wide and the second to fifth abdominal segments were membranous at the middle of the ventral surface, forming the so-called hunger folds. The primary genitalic structures of the female formed a recognizable homolog of the ovipositor and occupied the ventral aspects of the ninth and tenth segments.
Laterally, the paratergites of the eighth segment bore the last spiracles (Fig. 5). A pair of broad, flat plated called gonocoxae occupied the median two- thirds of the ventral aspect of the eighth segment. The lateral plates of the ninth segment were the fused paratergite and 2nd gonocoxa.
As shown in Fig. 5, a small incision was present on the right-handed posterior margin of the fifth sclerite. This incision is the opening of a copulatory pouch referred to as a spermalege (also known as paragental sinus or Berlese,s organ). Spermalege is an organ specialized to receive hypodermically injected semen. The spermalege consisted of two parts, the ectospermalege, and endospermalege. The ectospermalege derived from the ectoderm and consists of a groove. The mesospermalege is derived from mesoderm and is a membrane-bound sac attached to the wall of the haemocoel, directly beneath the groove of the ectospermalege [20].
In male bugs, the intromittent organ is highly specialized for traumatic insemination. During traumatic insemination, a male bug inserts his intromittent organ into the ectospermalege and pierces the pleural membrane. The sperm is then injected into the mesospermalege, which contains hemocytes [20]. The spermatozoa then form a mass and diffuse through the wall of the mesospermalege, swimming through the hemocoel to a pair of seminal conceptacles, where sperm are stored before they migrate to the ovaries for fertilization. Seminal conceptacles have evolved independently of the bursa copulatrix and spermatheca [20].
The paramere arose close to its apex on the ventral side. The paramere was strongly curved to the left, its external portion lying in a furrow in the side of the ninth segment. The base of the paramere was retracted into the body for a short distance. The paramere was grooved on the outer or convex side, except apically where it was folded to form a tube. The aedeagus lied just above the base of the paramere (Fig. 5). The aedeagus was a long narrow tubular structure whose walls were not sclerotized. When not in use the aedeagus was telescoped into the capsule-like phallobase. It was probably extended by fluid pressure.
The entire abdomen was covered with numerous chaetae on both dorsal and ventral sides. At the tip of the abdomen of both the male and female bugs, there presented a tuft of relatively long chaetae. Based on electron microscopic results we found that in female bugs chaetae had three teeth while males were with four teeth in the tip (Fig. 5).
{kind=link}
{kind=link}