Hyperactivity and inattention may be related to the insufficient function of the catecholamine (i.e., DOPA and NE) system in the brain. According to Cai et al., (2021), ADHD symptoms are associated with decreased synaptic DOPA and NE concentrations. In addition, there is a dysfunction of information transmission between inhibitory synapses and excitatory synapses (Xi, Wu et al., 2021), where occurs an atypical balance of neural excitation and inhibition (E/I) being GABA and glutamate as contributing (Mamiya et al., 2021). Although ADHD is associated with dysregulation of the catecholaminergic pathway and E/I balance in the brain, data also point to the contribution of oxidative stress and neuroinflammation in the pathophysiology of ADHD (Prince, 2008; Lopresti, 2015; Leffa et al., 2017; Darwish et al., 2019; Dursun et al., 2021).
SHR animals are used as ADHD models because they demonstrate hyperactive behavior even in familiar environments (Langen, Dost, 2011), anxiety (Ji et al., 2014), and spatial learning memory deficit (Kim et al., 2011). Disturbances in the functions of the neurotransmitters glutamate, DOPA, and NE in the brain of SHR and information obtained from patients with ADHD suggest a defect in neuronal circuits that are required for reward-oriented associative learning and memory formation (Russel, 2003). Sagvolden et al. (2005) suggest that the altered function of DOPA fails to adequately modulate the glutamatergic excitatory and GABAergic inhibitory signal transmission. Thus, persistent hypertension in SHR rats has depressive effects on glutamatergic and GABAergic parameters of the synaptic activity of neurons, which would influence the animals' learning and memory (Russel, 2003). The hypodopaminergic theory of ADHD states that hyperactive and inattentive behaviors are caused by low levels of DOPA, which leads to NMDA (N-methyl-D-aspartate) and AMPA (α-amino-3-hydroxy-5-methyl- receptors). 4-isoxazole propionic) become more active, resulting in increased glutamatergic production. In this situation, glutamate would normally release more DOPA, however, in the brain of individuals with ADHD, this feedback does not appear to occur (Miller et al., 2013). Furthermore, it is worth noting that the release of DOPA stimulated by glutamate in the nervous system of SHR is greater (Warton et al., 2009).
Furthermore, norepinephrine activity influences the acquisition and reinforcement of conditioned responses, and catecholamine-glutamate interactions strengthen and change the reinforced and adaptive behavior (Mingote et al, 2004). Therefore, the basic treatment for ADHD is stimulant drugs, eg, methylphenidate, which increases synaptic DOPA by directly blocking the DOPA transporter (DAT) (Biederman et al., 2004) or atomoxetine, which increases extracellular levels of noradrenaline (Arnsten, 2010). Also related to noradrenaline, in SHR, the inhibition of NE release mediated by the AMPA receptor, which is necessary for the NMDA effect, is impaired and glutamate stimulation of NE release is greater than that of Wistar Kyoto and Sprague rats –Dawley rats, in the prefrontal cortex and hippocampus (Howells, Russell, 2008).
The evaluation of several natural products has been carried out in search of possible therapies for ADHD. Thus, knowing that curcumin acts on several biological pathways including oxidative/nitrosative, and inflammatory (Wang et al., 2010; Bhandari, 2015; Tsuda, 2018; Pan et al., 2019), its protective actions are reported in several neurodegenerative and neuropsychiatric disorders (Mythri, Bharath, 2012). This compound can increase the inhibition of DOPA and NE reuptake by different pathways (Tabeshpour, et al., 2019), acting on neurotransmitter imbalances implicated in bipolar disorder (Anderson, Maes, 2015), anxiety disorders (Furtado, Katzman, 2015), schizophrenia (Davis et al., 2014) and autism (Rossignol, Frye, 2014).
Thus, the present study was designed by the fact that curcumin (30 mg/kg, p.o.) demonstrated neuroprotective action in diseases involving catecholaminergic deficits, mainly dopaminergic and noradrenergic deficits (Tamegart et al., 2019). Motawi et al. (2020) demonstrated that curcumin (80 mg/kg/day, p.o.) can significantly increase noradrenaline levels in the brain of rotenone-induced Parkinson's disease rats. In the same model, Madiha, Haider (2019) found that curcumin (100 mg/kg p.o.) for 2 weeks improved neurotransmitter levels as compared to rotenone injected rats. In a depression model induced by reserpine, curcumin-coated iron oxide nanoparticles for two weeks elevated serotonin (5-HT), NE, and DA levels (Khadrawy et al., 2021). Saied et al. (2020) demonstrated the same result, where curcumin (100 mg/kg, p.o., daily for one month) improved DOPA and NE levels in striatum and hippocampus in ovariectomized rats. They also showed that the treatment resulted in the down-regulation of monoamine oxidase b and up-regulation of tyrosine hydroxylase, as well as the DOPA receptor mRNA in the limbic region. In addition, inflammatory and oxidative parameters were ameliorated in the limbic system.
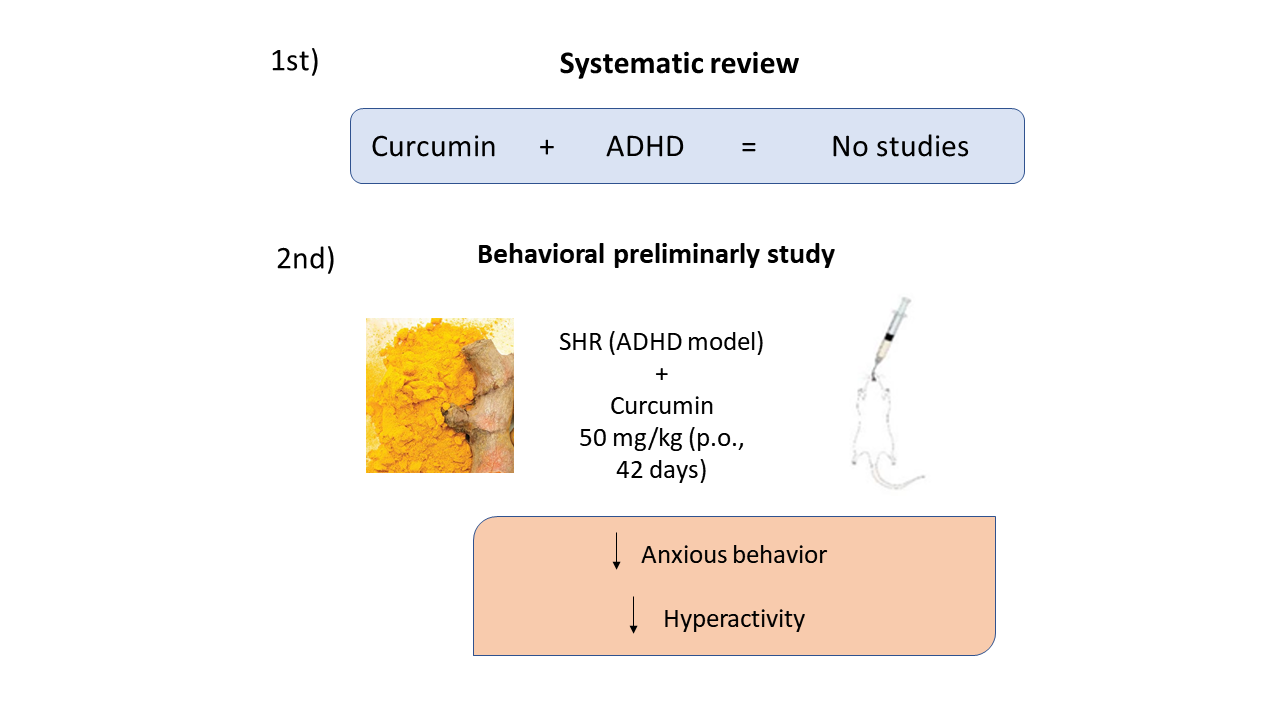
In this study, animals SHR treated with vehicle (SHR-V, negative control) moved around and further explored the open field arena. On the other hand, SHR animals treated with curcumin (SHR-C, 50 mg/kg/day, p.o.) for 42 days tended to demonstrate less anxious and hyperactive behavior, identified by the number of crossings in the open field test and active behavior in the social interaction test. Methylphenidate-treated animals also showed differences in locomotor but not exploratory behavior compared to SHR-V. Although anxious behavior measured by the EPM was not modulated by any treatment, Kishikawa et al., (2014) state that this test also reflects SHR's impulsiveness in search of novelties, which we can also suggest was unchanged. In the social interaction test, curcumin significantly showed more active behavior. In this test, environmental manipulations (lighting and familiar or unfamiliar arena) can increase or decrease the number of times rats interact, thus allowing the assessment of anxiolytic (increased interaction) or anxiogenic (decreased interaction) effects (Gogas et al. al., 2007). In this sense, the anxiolytic activity of curcumin has already been determined in several studies (Ceremuga et al., 2017; Lee, Lee, 2018; Marques et al., 2021).
In a recent systematic review, Sanei and Saberi-Demneh (2019) revealed that curcumin moderated or reversed the memory deficit in animal models in rodents, not being considered a placebo effect. However, the results of the Y-maze tests in our study demonstrate significant differences where SHR-C animals entered more and spent more time exploring the new arm of the apparatus when compared to SHR-M and KYT-V animals. However, in the object recognition test, only SHR-M affected long-term memory. Thus, the results are in line with those of Guo et al. (2012), where chronic methylphenidate administration did not affect the spatial learning performance of SHR animals during Morris water maze training, but significantly improved memory. According to the systematic review by Leffa et al. (2019), methylphenidate increases attentional and mnemonic performance in SHR animals. However, the drug does not reduce hyperactivity in SHR at low and medium doses. It should be noted here that according to Sontag et al. (2013), data on spatial working memory deficits in SHRs are not consistent, as they may be related to locomotor activity. Furthermore, the authors also mention that compared to KY rats, SHRs did not show any impairment in spatial working memory and reference memory. When the locomotor activity of rats was taken into account, the working memory and reference memory of SHRs were significantly better than in KY rats, according to the authors.
Many studies are carried out concerning the neuroprotective aspects of curcumin, especially in neurodegenerative diseases. However, related to ADHD, no study was developed and completed. It should be noted that this review was restricted to Portuguese and English, therefore studies in other languages may have been ignored in the selection of papers. Although we have not found any preclinical or published clinical studies linking ADHD with curcumin, this review suggests that studies should be continued to clarify the possible use of curcumin as a therapeutic agent.
Based on the above, the available literature on the effects of curcumin in SHR animals with a focus on neural aspects remains empty. Therefore, through the results of the present study, it can be suggested that this non-pharmacological approach could lead to beneficial effects on behavior in SHR. However, it is worth noting that there is a limitation of the low statistical power of the sample size. Therefore, studies should be conducted in order to assess the time and dose-dependent effect of curcumin treatment and its association with longer exercise protocols in SHR animals. Higher concentrations of curcumin could be needed to produce a more pronounced effect given its low oral bioavailability (Lopresti, 2018). However, in this study, the oral route was chosen as it is the most routine route of administration in clinical practice. Although with limitations, the findings suggest that the protocols used in this study are of great value to modulate the behavior of these animals, but further investigations are needed.
{kind=link}