Maize photosynthetic performances variations in response to drought stress
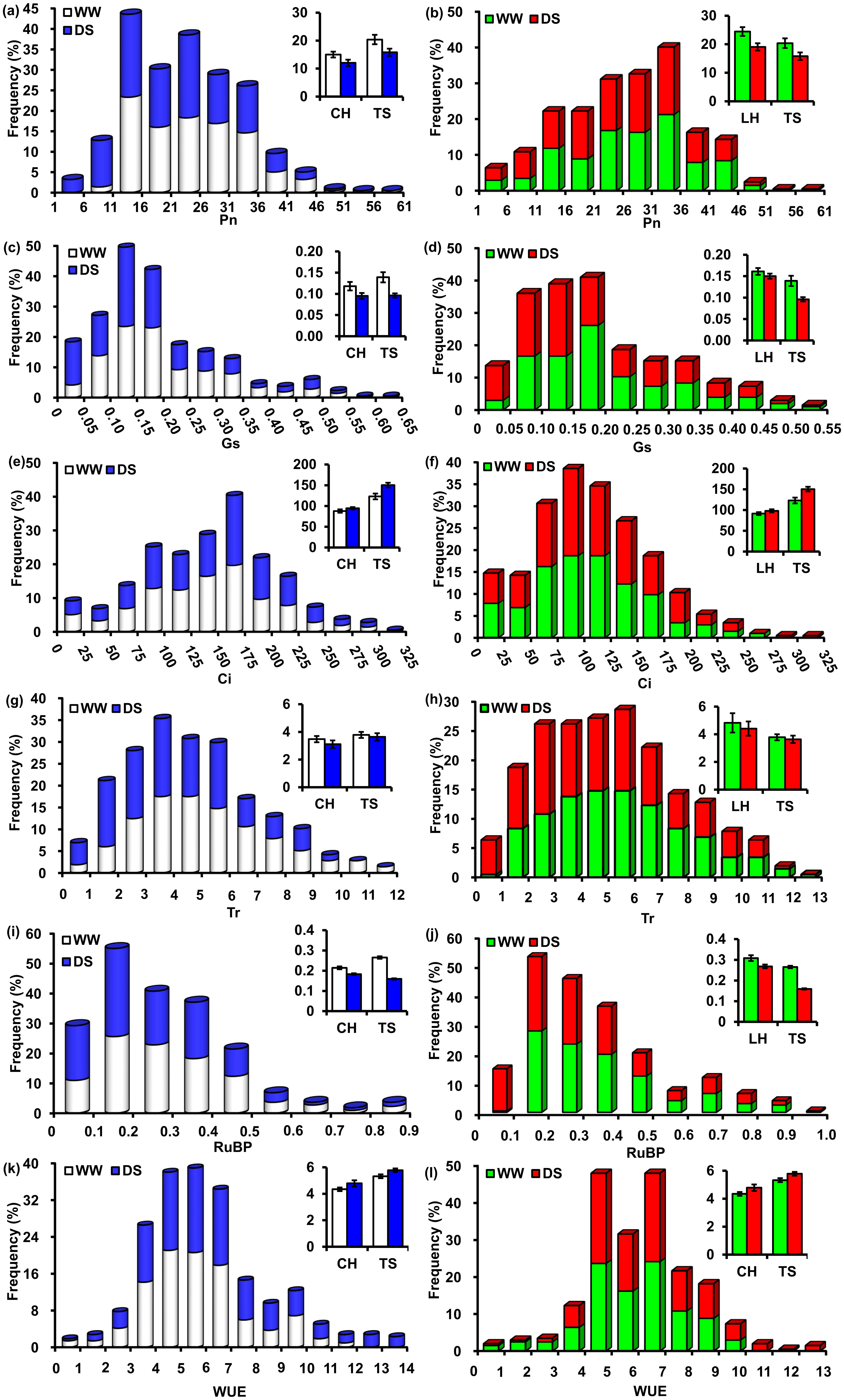
Approximately 95 % of the organic matter accumulated by crops comes from its own photosynthesis, and photosynthesis efficiency could directly determine the GY. Drought stress can significantly affect crop photsynthesis, and the responses of leaf photosynthesis to drought were mediated by two different physiological processes [35-36]. Firstly, SC and AMC decrease were recognized as the main driver of the photosynthetic response to drought stress, in order to reduce transpiration under water deprivations, the plant stomatal can close, and SC could limite CO2 diffusion efficiency from the atmosphere to the substomatal cavities to slow photosynthesis [37-38], and AMC rapidly decrease could further limite CO2 diffusion efficiency from the substomatal cavities to the chloroplast stroma during water stress [39-40]. Secondly, photosynthesis could be limite by biochemical processes resulting in photosynthetic enzyme activity inhibition, Pn, Rubisco and RuBP activity decrease, etc. [7-9, 36]. Fortunately, above these findings were also supported by our results in this study, namely, drought-stressed Chang7-2, Langhuang, TS141, POP-CT, and POP-LT exhibited obvious decrease in Pn, Gs, Tr, and RuBP, their Ci and WUE, however, significantly displayed the increase relative of corresponding plants with sufficient water (Fig. 1, Additional file 2: Table S1). Furthermore, RuBP (average RC 18.96 %) and Gs (average RC 17.05 %) in response to drought stress were more sensitive than the response to Pn (average RC 16.02 %), Ci (average RC -9.34%), Tr (average RC 10.80 %), and WUE (average RC -8.62 %) (Fig. 1). It could be concluded that the drought-induced limitation of photosynthesis in maize was primarily due to CO2 diffusion efficiency from sub-stomatal interval internal cavities to carboxylation site in chloroplasts and degree of SC, and increasing evidences in maize also supported our results by Liu et al. [5],Veroneze-Júnior et al. [41], Perdomo et al. [42], and He et al. [4]. Therefore, genetic improvement of photosynthetic performances in maize can be applied to MAS breeding to improve drought tolerance and high-yielding in the future.
Genetic architectures for photosynthetic performances
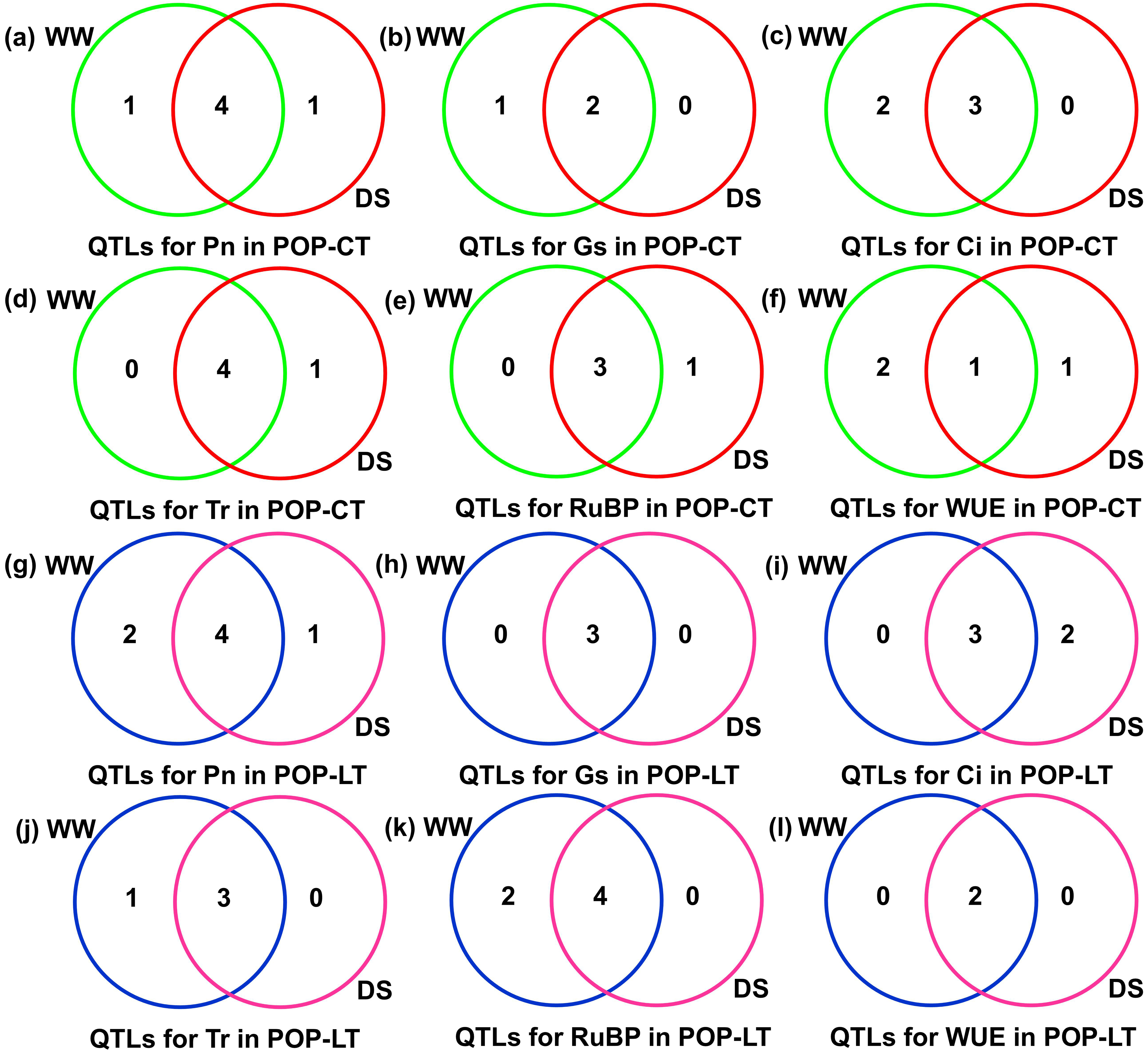
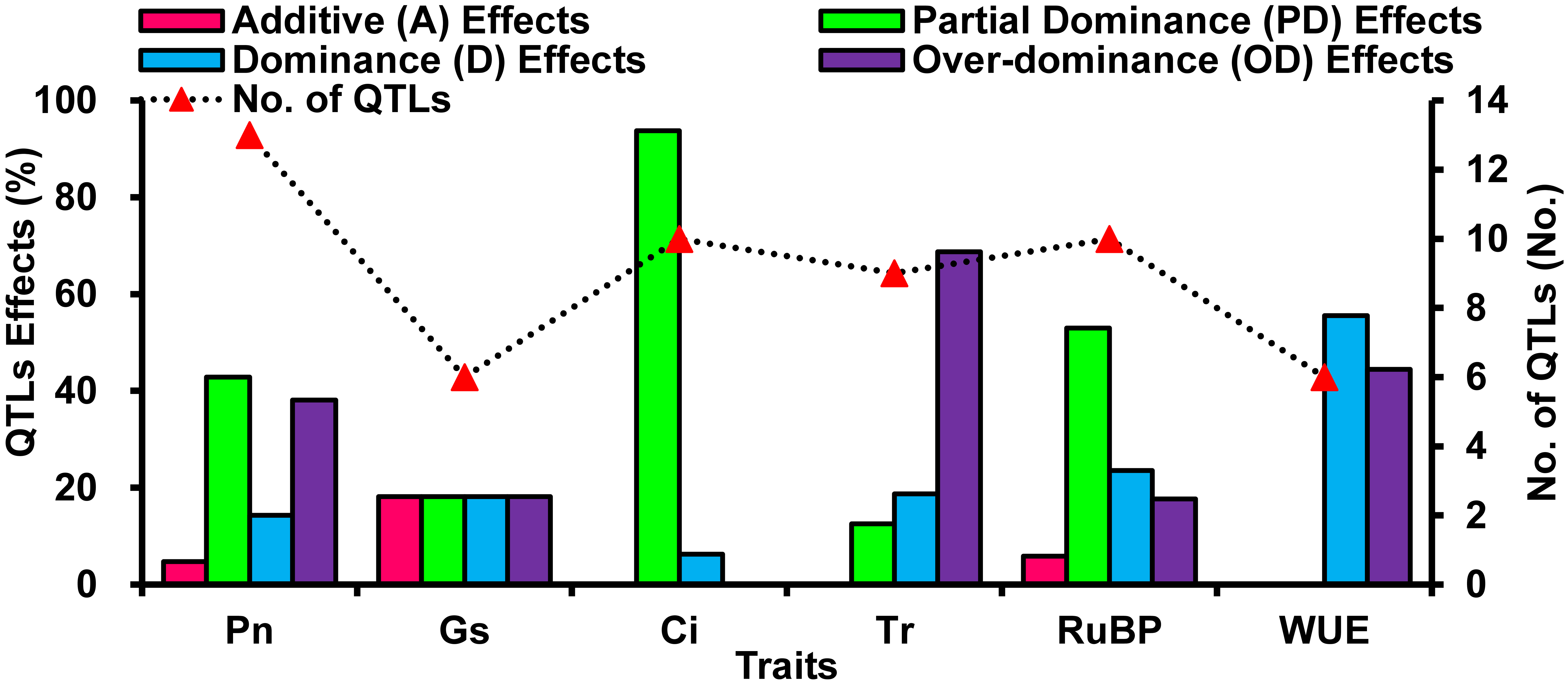
Although a wealth of information from previous researches considerably improved our understanding of leaf photosynthetic performances [43-45], as well as applications in maize MAS breeding [5, 9, 18, 35], few studies considered the genetic basis of maize photosynthetic-related traits under water defict at the molecular level [20-22]. Based on the above considerations, in this study we detected 54 QTLs for six photosynthetic-related traits across two F4 populations via single watering environment mapping with CIM (Fig. 4, Additional file 6: Table S3), and for the identified QTLs, Pn, Gs, and RuBP showed both additive and non-additive effects under drought and non-drought stressed environments, but non-additive effects were largely responsible for the genetc basis of these three traits, of which accounted for 95.24, 81.82, and 94.12 %, respectively (Additional file 5: Fig. S3). However, all identified QTLs for Ci, Tr, and WUE displayed non-additive effects under both contrasting watering environments (Additional file 5: Fig. S3). These results were consistent with Wang and Zhang [18] and Li et al. [46] involved in photosynthetic-related traits in maize. Further Pearson correlation analysis among F4 progencies and their parents showed that Pn, Gs, Ci, RuBP, and WUE of the two F4 populations depicted significantly positive correlation to female parent, as well as Tr showed significantly positive correlation to both parents (Additional file 4: Table S2). Thereby, breeders should pay more attentation to the evaluation of F1 cross combinatons to make good use of their prominent non-additive effects and specific combining ability for above six photosynthetic-related traits, as well as should carefully select parents with elite photosynthetic-related traits, finally achieving the aim of improving these traits under drought and un-stressed environments. Additionally, 43 of the 54 identified QTLs were found in drought-stressed environments (Additional file 8: Fig. S4). Suggesting that these identified QTLs controlling Pn, Gs, Ci, Tr, RuBP, and WUE could be changes under both contrasting environments, and the QTLs were identified under drought stress that may directly lead to differences in Pn, Gs, Ci, Tr, RuBP, and WUE.
GEI is critical in determining the adatation and fitness of genotypes in adverse environments, resulting in phenotypic variations [47-48], QEI information thus obtained was of great value for breeders and genetic researchers [47]. Furthermore, the variations of photosynthetic-related traits in two F4 populations showed that the wide variations, i.e. , , and were observed for six photosynthetic-related traits (P < 0.01 or P < 0.05), as well as their ranged from 1.466 % to 19.770 % (Table 1). Further analysis of QEI in two F4 populations with MCIM among all watering environments also suggested that totally 24 QEIs (44.44 %) of the identified joint QTLs controlling Pn, Gs, Ci, Tr, RuBP, and WUE were mapped, and each QEI explained 1.34~6.86 % of the phenotypic variance by h2(AE) in present study (Additional file 8: Table S4). In this regard, photosynthetic-related traits are similar to other traits, such as yield-related, and leaf and inflorecence architecture traits in maize [27-29], of which showed extensive GEI. As a result, GEI may be a major challenge to MAS breeding for photosynthetic-related traits in maize.
Epistasis, i.e. the interactions between genetic loci, is also thought to contribute to photosynthetic performances variations [49-51]. In maize, Li et al. [46] reported that SPAD was controlled by 2 major genes with AD-epistatic effects and polygene with AD effects by mixed major gene plus polygene genetic model. Liu et al. [52] also suggested that SPAD was in agreement with the AD-epistatic model using diallel cross II (Griffing). As expected, in accord with previous studies [46, 52-53], totally 14 pairs of epistatic interactions with DA and DA controlling Pn, Gs, Ci, Tr, RuBP, and WUE based on different watering environments in two F4 populations, and which accounted for 2.09~5.87 % of the observed phenotypic variance contributed by h2(DA/DD) that were clearly lower than those from A effects for all photosynthetic-related traits (Table 3, Fig. 5). It could be concluded that low contributions to phenotypic variance explained by DA/DD effects were due to large No. of DA/DD-QTLs with minor genetic effects, which would significantly influence the efficiency of MAS breeding for photosynrhetic-related traits. Notably, two stable DD-epistatic interactions were also validated in our study, i.e., controlling Pn between bin 1.07_1.08_1.10 (bnlg1025-mmc0041-phi308707) and bin 10.03 (bnlg1655-umc2016/umc1345) was repeatedly detected in POP-CT under both contrasting watering environments/POP-LT under stressed environment, and affecting Ci between bin 1.08_1.10 (mmc0041-phi308707) and bin 6.05 (umc2040-bnlg1174a) in two F4 populations under both all four environments (Table 3, Fig. 5). These results were supported by Zhao et al. [26] and Sa et al. [54], who indicated that an AD/additive-additive (AA) epistatic interaction for KW, plant height, and ear length, etc., located on Chr. 1 and Chr. 6/10 under multiple drought and non-drought environments. Hence, these regions of chromosomes may be considered as epistatic regulators that influence maize plant development, photosynthesis, and ear formation under multiple watering regimes.
cQTLs comparison and candidate genes validation for photosynthetic performances
The identified cQTLs for photosynthetic-related traits in a broad genetic background under adverse watering environments could provide guidance for fine mapping and maize MAS in the future. We totally identified 8 cQTLs for six photosynthetic-related traits using CIM/MCIM across POP-CT and POP-LT under multiple watering environments, of which accounting for 3.21~15.78 % of the average observed phenotypic variance in this study (Table 2).
Af these, cQTL2 in bin 1.07_1.10 (bnlg1025/mmc0041-phi308707/umc1847) was simultaneously associated with Pn, Ci, Tr, and WUE, sQTL5 in bin 6.05 (umc2141-umc2040-bnlg1174a) was simultaneously involved in Pn, Tr, Ci, and RuBP, sQTL6 in bin 7.02_7.04 (umc2057-bnlg1666-umc1708) was simultaneously affected Pn and RuBP, sQTL7 in bin 8.03 (bnlg1863-umc2075) was simultaneously responsile for Ci, Tr, and WUE, and sQTL8 in bin 10.03 (bnlg1655-umc2016/umc1345) was simultaneously associated with Pn , Tr, and RuBP in two F4 populations under both constracting watering environments, suggestive of pleiotropic cQTLs, which were highly agreed with the Pearson correlations analysis among six photosynthetic-related traits under both watering environments (Fig. 3). In bin 1.08_1.10 (umc83a-umc39c), in bin 6.05 (near gsy298e_pmg), bin 7.03_7.04 (gsy113_cs-gsy107_pr), and bin 10.03 (gsy321_aba-gsy329_pp) intervals, Pelleschi et al. [21] also detected multiple QTLs associated with photosynthesis (i.e., sucrose-P synthase (SPS), hexoses (HEX), sucrose (SUC), AGP, Tr, and CO) and leaf morphologies (i.e., number (LN), width (LW), length (LL), and relative water content (RWC)) across 120 F-2 × MBS847 RILs under drought and non-drought conditions. Li et al. [50] also mapped multiple QTLs involved in SPAD in bin 1.08 (umc1013-umc2047), 7.02 (umc1585-bnlg1305), bin 8.03 (bnlg1863-bnlg2046/umc2075-phi100175), and bin 10.03 (bnlg1655) regions in 172 Xu172 × Zong3 single segment substitution lines (SSSLs) under both high and low N treatments. Wang and Zhang [18] also identified two pleiotropic QTLs were simultaneously responsible for FCa, FCb, and FCt in bin 1.08 (mmc0041-bnlg1556) and bin 8.03 (umc1457-umc2199) using 189 A150-3-2 × Mo17 F2 plants under a single environment. Peng et al. [33] also found a QTL for grain tield per plant (GYPP) in near phi308707 (bin 1.10), a stable QTL controlled GYPP and kernel number per plant (KNPP) located in bnlg1094-bnlg1579 (bin 7.02_7.03), and a QTL affecting KW in bin 8.03 (bnlg1352-umc1778) cross 230 Qi319 × Huangzaosi and 235 Ye478 × Huangzaosi F2:3 families during six environments. These results indidated that pleiotropic cQTLs in the bin 1.07_1.10, 6.05, 7.02_7.04, 8.03, and 10.03 regions may play critical roles in leaf development, photosysthesis, and yield formation in maize under contrasting watering environments, and some important genes may be located in these bin regions. As expected, totally 11 cnadidate genes were also validated in above 5 pleiotropic cQTLs (cQTL 2, cQTL5, cQTL6, cQTL7, and cQTL8) intervals. GRMZM2G018627 (LHCB9) is a light harvesting chlorophyll binding (LHCB) protein, and Zhao et al. [55] proved that LHCB protein was required for the maintenance of photosystem I and specific protein-chlorophyll complexes especially under certain stress conditions. GRMZM2G162672 (chlg1), i.e., chlorophyll synthase G1 gene [56], acted as a chlorophyll biosynthetic process in biological progress via the AgBase v. 2.00 (http://agbase.arizona.edu/) online software with gene ontology (GO) analysis. GRMZM2G039113 (tan1), i.e., tangled 1 gene was required for spatial control of cytoskeletal arrays associated with cell division during maize leaf development [57], as well as the tan1 mutation could alter cell division orientations throught leaf development without altering leaf shape, and even affected maize photosynthesis capacity [58]. GRMZM2G013657 (dwil1), i.e., dwarf & irregular leaf 1 gene may regulate leaf base, tip, and sheath development in maize (https://maizegdb.org/ gene_center/gene). GRMZM5G809292 (PYG7), i.e., tetratricopeptide repeat domain-containing protein PYG7 [59], was the components of the chloroplast and thylakoid membrane in mellular component and involved in photosystem I assembly in biological progres by GO analysis. GRMZM2G042592, encoded the thioredoxin-like 6, chloroplast contained a variety of thioredoxin systems [60], while the thioredoxin could interact with CHLI subunits of Mg2+ chelatase to regulate the chelation of Mg2+ chelatase and chlorophyll synthesis precursor protoporphyrin IX [61]. Wang et al. [62] also predicted GRMZM2G042592 located in the same region of bnlg1863-umc2075 (bin 8.03), and its mutant with abnormal chloroplast, lacking pigment and reducing of PSII. GRMZM2G163437 (agps11), i.e., ADP glucose pyrophosphorylase small subunit leaf 1 gene, could be expressed in maize grain and source leaf during grain filling [63]. GRMZM2G033885 (psb29), encoded photosystem II subunit29, which was reversibly phosphorylated in maize upon exposure to high light in the cold condition, as well as its phosphorylation was controlled by the redox state of the plastoquinone pool, and may be part of a novel mechanism of photoprotection [64]. Another 2 candidate gens, i.e., GRMZM2G045431 (bHLH150) and GRMZM2G058451 (bHLH164), belonging to bHLH transcription factors, which played important roles in maize grwoth and development, such as, root differentation [65], photomorphogenesis and light signal transduction [66], and stress response [67].
In addition, We also noted that cQTL3 affecting Gs in bin 3.07_3.08 (umc1286/umc2275-umc2081) with CIM/MCIM cross two F4 populations under drought and non-drought environments. Li et al. [50] also found a QEI for SPAD in bin 3.08 (umc1844-bnlg1182) under high N condition, and even Guo et al. [68] reported a meta-QTL (mQTL) for LW in bin 3.08 region from 28 original populations under multiple environments via mQTL analysis. Then further analysis validated two candidate genes, i.e., GRMZM2G159937 (bHIH57) and GRMZM2G117851 (bZIP1) in the bin 3.07_3.08 intervals in our study. GRMZM2G159937 may have all the functions of bHLH transcription factor family [65-67]. Walsh et al. [69] reported liguleless2 (lg2) gene encoded a bZIP protein, involved in maize ligule and auricle development of leaf during vegetative and reproductive growth periods. GRMZM2G117851 was also a bZIP transcription factor, may play an important role in maize leaf development.
Additionally, 2 new cQTLs (cQTL1 and cQTL4) information were also found in our study. cQTL1 controlling WUE in bin 1.00_1.01 (bnlg149-bmc1014/umc1177) with CIM/MCIM cross both populations, of which GRMZM2G042250 (rld2, rolled leaf 2) was predicted, and its orthologous gene ATHB23 (i.e. a phytochrome B-interacting protein), was important for phytochrome B-mediated red light signaling in Arabidopsis thaliana [70]. cQTL4 responble for Ci in bin 4.08_4.09 (umc2041-umc2188/umc2287) with CIM cross both populations under stressed and un-stressed environments. GRMZM2G446426 (MADS52), GRMZM2G038479 (bHLH8), and GRMZM2G074122 (pep3) were identified in cQTL4. GRMZM2G446426 as the MADS-transcription factor 52, could have the similar as ZmMADS4 in influencing chlorophyll content, and involve in response to osmotic stress [71]. GRMZM2G074122 was phosphoenolpyruvate carboxylase isoform 1 gene, which involved in carbon fixation and tricarboxylic acid cycle in biological process and influenced phosphoenolpyruvate carboxylase activity in molecular function via GO analysis. As a result, the 2 cQTLs intervals may provide new information for genetic basis dissection in maize photosynthetic performances under both watering environments in the future.
{kind=link}
{kind=link}
{kind=link}
{kind=link}