An ex-vivo kidney perfusion system for evaluation of xenograft rejection
Before transplanting a xenograft kidney in patients, it is essential to evaluate its safety and efficacy in preclinical human-like conditions. To realize this, we established an ex vivo normothermic pig kidney perfusion system as shown in Fig. 1A. The system was adapted from clinically used systems. We collected kidneys from one wildtype (WT) pig and one genetically modified pig (4KO) lacking three xenoantigens: GGTA1 encoding the Alpha-1,3-galactosyltransferase, CMAH encoding the cytidine monophospho-N-acetylneuraminic acid hydroxylase, and B4GALNT2 encoding the Beta-1,4-N-Acetyl-Galactosaminyltransferase 2, as well as lacking the swine leukocyte antigen (SLA) class I molecule (SLA-I). The kidneys were perfused with either pig blood (PB) or human blood (HB). Pressure regulated flow supplied the renal artery with carbogen oxygenated whole blood at 37 ˚C (Fig. 1A and Supplementary Table 1). Perfusion of the WT kidney with HB resulted in acute rejection and the perfusion could not be sustained longer than for six hours because of clotting activation. Medulla and cortex biopsy samples from WT and 4KO kidneys were histologically examined and analyzed by H&E staining to study signs of rejection.
Extensive renal injury was observed in all renal compartments from WT pig kidneys perfused with HB, while in the 4KO kidney this injury was strongly reduced (Fig. 1B). To quantify the renal injury, H&E images (n=3) from biopsies were scored by an independent and certified pathologist. 14 renal injury parameters were scored using the Endothelial, Glomerular, Tubular, and Interstitial (EGTI) components-based comprehensive histology scoring system19, 20. Intratubular hyaline, tubule epithelial cell necrosis and degeneration, interstitium hemorrhage, glomeruli microthrombi and hypercellularity were significant (t-test, p value < 0.005) reduced in the 4KO compared to the WT (Fig. 1C). Particularly, there is less tubular injury in both cortex and medulla in the 4KO than the WT. Broad and strong microthrombosis was observed in the WT glomeruli perfused with HB, which is significantly (t-test, p < 0.005) reduced in the 4KO pigs.
Elimination of pig xenoantigens has been shown with benefits on prevention of immune activation in transplantation experiments 3, 21–23. To validate this in our perfusion model, we stained the perfused kidneys tissues with antibodies against human CD3 and CD4, which are markers of T lymphocytes. We observed strong accumulation of T lymphocytes in the renal cortex of WT pigs perfused with human blood, particularly in the glomeruli, whereas the CD3 or CD4 staining signal in the 4KO kidneys perfused with human blood is strongly reduced (Fig. 1D). CD3 and CD4 is colocalized at the endothelium as validated by co-staining the tissues with endothelial cell marker CD31, indicating that there is an accumulation and infiltration of human T lymphocytes at the endothelium in the WT-HB pig kidney. In addition, we performed complement activation staining on the WT-HB and 4KO-HB tissues. The WT-HB shows C4d and C5b9 positivity mainly within the glomeruli pointing towards activation of the complement system whereas this has disappeared in the 4KO-HB (Fig. 1E). Both coagulation and complement activation have been known as the key cascades in mediated the organ rejection 24, 25. As a further validation of coagulation, after biopsy sampling, we recalcified the perfused blood re-establishing a physiological coagulation cascade within the perfusion system. Mean arterial pressure was measured in time showing a significant increase in the WT kidney perfused with human blood compared to a moderate increase of the 4KO (Fig. 1F), suggesting that genetic elimination of the SLA-I and three xenoantigens significantly reduces xenograft-rejection associated coagulation. Collectively, we generate an ex vivo system to evaluate xenograft rejection in a preclinical human-like condition and validate that elimination of xenoantigens can significantly reduce the acute renal xenograft rejection-associated kidney injuries.
Single cell RNA sequencing of renal endothelial cells
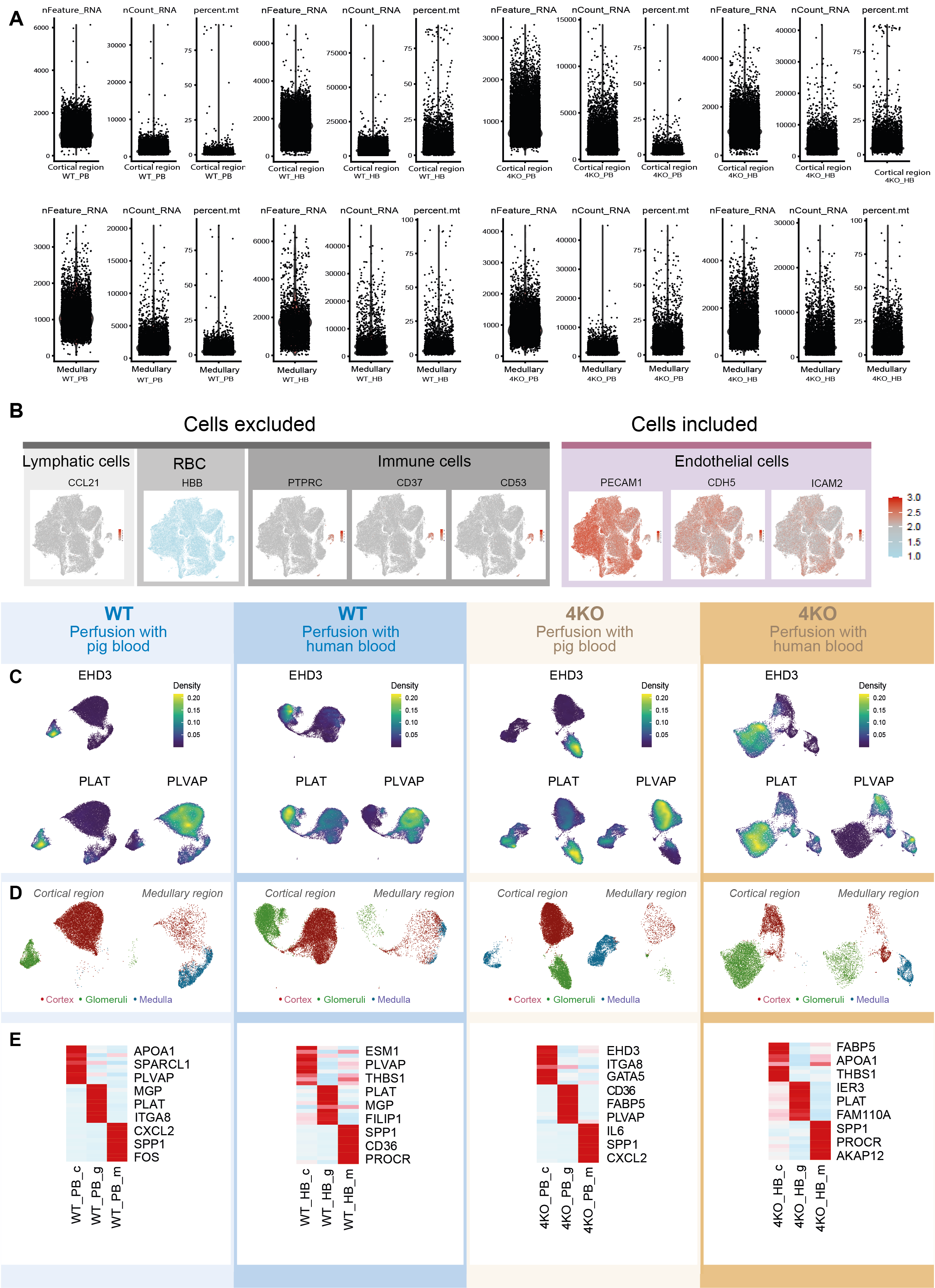
Blood endothelial cells play important roles in the modulation of xenograft rejection processes. To gain deeper insights into xenograft rejection of renal endothelium and the alteration of molecular signatures of endothelial cells, we isolated renal endothelial cells (REC) from biopsies of perfused kidneys at six hours and analyzed the endothelial cell transcriptome by single cell RNA sequencing. Using a protocol of REC isolation established previously 18, single endothelial cell suspension was obtained by enzymatic digestion, followed by magnetic-activated cell sorting (MACS) and fluorescence-activated cell sorting (FACS)-based enrichment of ECs expressing CD31. In total, we collected RECs from four groups: WT kidney perfused with pig blood (WT-PB), WT kidney perfused with human blood (WT-HB), 4KO kidney perfused with pig blood (4KO-PB) and 4KO kidney perfused with human blood (4KO-HB) (Supplementary fig. S1). In each group, single cell RNA sequencing was performed for RECs isolated from cortical and medullary biopsies separately (see methods). To avoid the freeze-thaw caused cell death, sorted cells were immediately subjected to scRNA-seq library construction using the Gel Bead-in-Emulsion (GEM)-based high throughput single cell RNA sequencing technology from 10X genomics, followed by deep sequencing (Fig. 2A). We obtained a sequencing saturation above 70% in all samples, with an average of over 50,000 reads per cells. In total, we captured single cell transcriptome from 93,988 cells. Based on the annotation of pig genome assembly Sscrofa11.1, we detected an average of 1136 transcribed genes per cell and 16,244 expressed genes per samples (Supplementary Table S2). Among the 93,988 cells, 2043 cells were removed due to either potential doublets or low quality (see methods). We further performed a marker-based silicon selection to exclude lymphatic cells (CCL2126), red blood cell (HBB), and immune cells (PTPRC, CD37(a marker for B cells)27, and CD53 (a thymocyte marker)28). Only cells expressing endothelial cell markers PECAM1 (CD31, a pan endothelial cell marker), CDH5 (VE-cadherin, blood vessel marker) 29 and ICAM2 (intercellular adhesion molecule 2 expressed between endothelial cell junctions) 30 were included for downstream analyses. After all these filtering steps, we obtained high-quality single cell RNA transcriptome from 70,140 RECs (38,519 from WT and 31,611 from 4KO) (Supplementary Table S3).
Previous single cell RNA sequencing of RECs in normal mouse kidneys showed that murine RECs exhibit compartmental specific transcriptome 18. To identify RECs from kidney cortex (cREC), glomeruli (gREC) and medullar (mREC), we applied two approaches to identify RECs from different kidney compartments: (1) Expression of glomeruli markers (PLAT and EDH3) and cortex/medulla marker PLVAP) 31; (2) Physical biopsy location to separate mRECs and cRECs/gRECs (Supplementary Figure S1). Our results showed that pig RECs exhibited distinct compartment specific transcriptome signatures (Supplementary Figure S1, Supplementary Table S4), consistent with previous observed of murine RECs 18. As expected, human blood perfusion strongly affects the REC transcriptome, as seen by both UMAP and correlation analyses (Fig. 2B-D). In WT and 4KO pig kidney, HB-perfused RECs were clustered together and were distinct from the PB-perfused cRECs and gRECs. Unlike cRECs and gRECs, the medullary endothelium (mRECs) displays a similar transcriptomic response to HB and PB. This indicates that HB induces a global transcriptome response (“the endothelial activation response”) in the glomerular and cortical endothelium (Fig. 2D). Although histological examination shows clear reduction of kidney injuries and signs of rejections by the elimination of SLA-I and three xenoantigens, single cell RNA sequencing suggests that human blood perfusion still has an effect on the global transcriptome changes in the 4KO pigs. To gain further insights into variation of pathway activities affected by human blood perfusion, we performed Gene Set Variation Analysis (GSVA) for 21 manually curated hallmark gene sets 32. Figure 2E summarizes endothelial transcriptome changes in a selection of pathways, where in particular changes in the interferon signaling pathways, which is characteristic of T cell mediated rejection, were reduced in the cortical and glomerular endothelial compartment of the 4KO. This was not a reflection of generalized inability to induce an endothelial transcriptomic response upon blood perfusion, as induction of e.g. inflammatory and hypoxia response pathways did not differ between the wild type and 4KO pigs. It should be noted that this analysis only reflects the gene expression variations, but not the activating or inhibitory effect of the corresponding pathways. Our results thus provide the first inventory of pig kidney endothelial cell transcriptome changes caused by human blood perfused at the single cell level and could serve to identify future targets to modulate rejection in pig xenograft transplantation.
Elimination of MHC class I (SLA-I) and three xenoantigens reduces renal endothelial transcriptome alteration caused by human blood perfusion
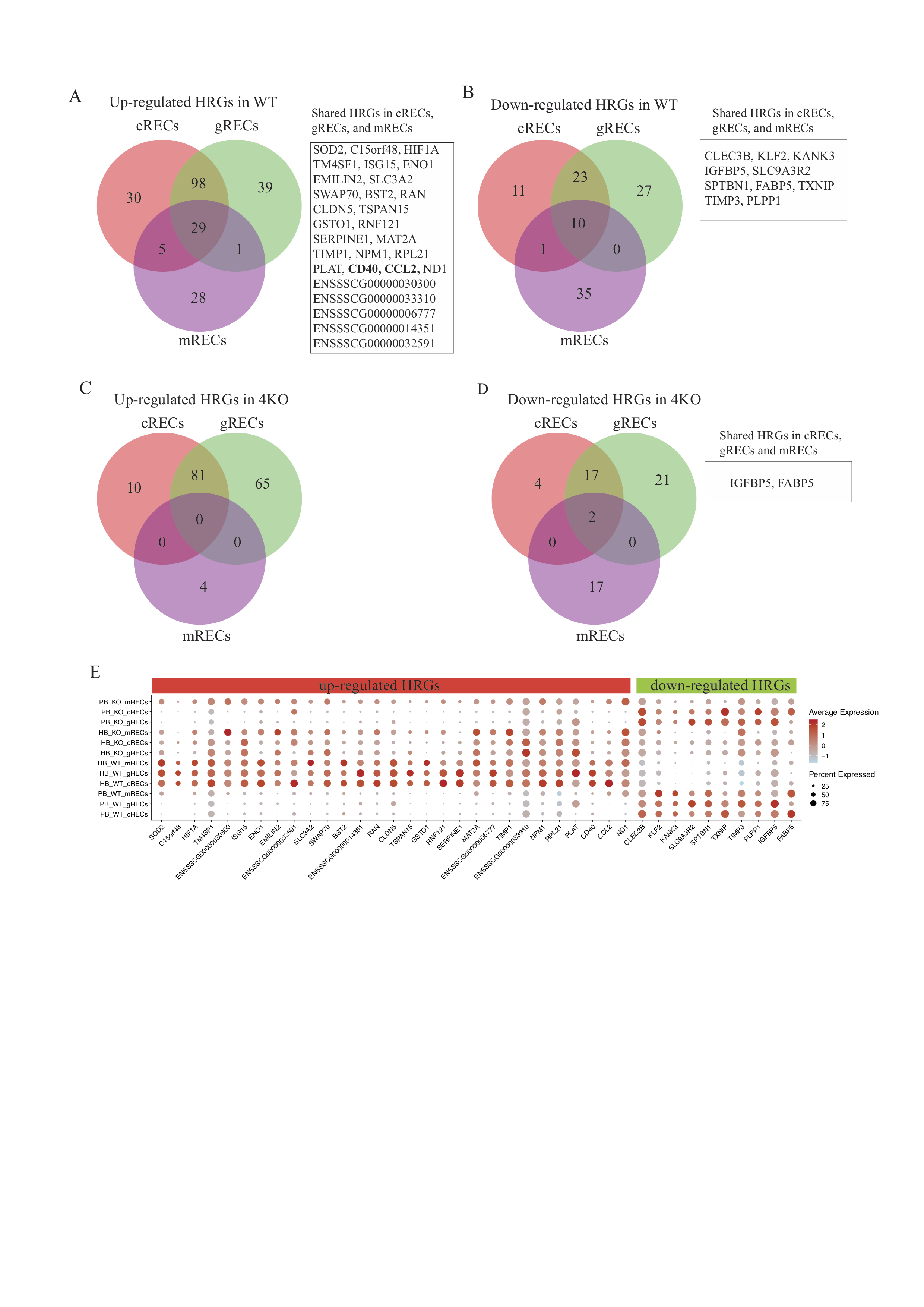
To further investigate the different response of WT and 4KO RECs to human blood perfusion, we compared cRECs, gRECs, mRECs, respectively, between HB and PB perfused kidneys. Differentially expressed genes (DEGs) were classified into three classes: High response genes (HRGs, absolute (abs.) log2 fold changes (FC) >= 1, adj. P < 0.05), low response genes (LRGs, 0.25 < abs.log2FC < 1, adj. P < 0.05), and not significantly responding genes (NRG). In WT pigs (Fig. 3A), we identified 1064 up regulated (162 HRGs and 901 LRGs) and 519 down regulated (45 HRGs and 474 LRGs) DEGs in cRECs, 1057 up regulated (167 HRGs and 890 LRGs) and 668 down regulated (60 HRGs and 698 LRGs) DEGs in cRECs, 1055 up regulated (63 HRGs and 992 LRGs) and 406 down regulated (46 HRGs and 360 LRGs) DEGs in mRECs. Although the number of up regulated DEGs is similar between cRECs, gRECs and mRECs in the WT pig, there are more than three-fold less HRGs in the mRECs compared to the cRECs and gRECs. The total number of down regulated DEGs are less (more than 100 genes) in mRECs compared to cRECs and gRECs. The differential gene expression analysis further suggests that HB perfusion triggers higher endothelial transcriptome activation in the cortical regions. Compared to the WT pig, the numbers of DEGs (both HRGs and LRGs) in cRECs, gRECs and mRECs of the 4KO are significantly lower (Fig. 3A, Supplementary Table S5). Approximately half of the LRGs in the WT RECs are NRGs in the 4KO RECs. Interestingly, our analysis revealed that approximately half of the HRGs from the WT RECs were LRGs in the 4KO RECs. These findings suggest that elimination of the four genes has substantially (although not completely) reduced HB-triggered endothelium transcriptome activations. To investigate what biological functions these DEGs are mainly involved in, we focus on the HRGs reasoning that the high response genes resembles the strong effects and provide more solid insights into the major effect on endothelial cell transcriptome by the elimination of SLA-I and the three xenoantigens. As shown in Fig. 3B and fig. S2, in WT pigs, there were 162, 167 and 63 up-regulated HRGs found in cortex, glomeruli and medulla respectively. In the 4KO pigs, there are less up-regulated HRGs (91, 146 and 4 found in cortex, glomeruli and medulla respectively). There are 65, 95 and 2 commonly up-regulated HRGs in both WT and 4KO kidney in cortex, glomeruli and medulla, respectively. Similarly, there are less down-regulated HRGs found in the 4KO RECs compared to WT RECs (Fig. 3B).
To characterize the genes actively responding to human blood perfusion, we performed gene ontology (GO) analysis for the up-regulated HRGs found in WT only (blue), common (gray), and 4KO only (orange) in all three compartments. Showed in Fig. 3C, the WT only HRGs were enriched in immune related pathways such as regulation of I-κB kinase/NF-κB signaling, toll-like receptor signaling pathway and positive regulation of interleukin-12 production, suggesting that these WT only HRGs are involved in active immune responses. The common HRGs were enriched in functions related to apoptotic process, cell death and would healing. Only the 4KO HRGs were found to be enriched in pathways related to ribosomal assembly and splicing, indicating an effect on the RNA processing machinery. The expression levels for genes commonly involved in these enriched pathways of cortex, glomeruli and medulla RECs were further displayed in the dot plot (Fig. 3D). C–C motif chemokine ligand 2 encoding gene CCL2 and member of the TNF-receptor superfamily gene CD40, which are involved in the regulation of EC apoptosis, inflammatory response, and immune system process, are highly up regulated in the HB-perfused WT RECs but greatly diminished in the 4KO. Genes involved in regulating the NF-kB signaling (i.e. Toll-like receptor 4 encoding gene TLR4 and TNF-α-induced protein 3-interacting protein 1 encoding gene TNIP1), which were more strongly activated in the HB-perfused WT gRECs, were also diminished in the 4KO. Immunostaining of TLR4, CCL2 and CD40 further confirmed upregulation at the protein level in the HB-perfused WT RECs but not the 4KO RECs (Fig. 3E). Together, our results suggest that human blood perfusion activates the expression of immune regulating genes of WT porcine renal endothelial cells, which was substantially diminished in the 4KO pigs.
Human blood perfusion activates proteasome complex and NF-κB pathway
To further understand the immunoregulatory roles of endothelial cells in xenograft rejection, we generated a subset of immune related genes (717 genes) from the entire dataset. This selected set was subjected to further DE gene analysis (abs.log2FC > 0.25, adjust P < 0.05) and were likewise grouped to WT only DEGs (blue), common DEGs (gray) and 4KO only DEGs (orange). More DEGs genes were up-regulated in all three compartments, indicating that perfusion with human blood led to activation of immune related genes (Supplementary Table S6). Within all three compartments, the WT kidney showed the largest group of upregulated unique genes (Fig. 4A). The heatmap in Fig. 4B shows the expression levels of the top selected genes in each group. Within the WT kidney many immune related genes are highly expressed (i.e. TAP1, ICAM2 and VCAM1), while in the KO kidney these genes show a low expression. Our results also showed that there is still a large group of commonly expressed immune genes, indicating a downregulation of the immune response by the 4KO model, but not a complete elimination.
In order to find differences and similarities among different compartments/genotype, we used upSetR to visualize the top gene sets which contain either shared or unique genes (Fig. 4C). Interestingly, the largest gene set, which contains 32 genes (see detailed gene list in supplementary table S6), was upregulated in all three WT compartments but not the 4KO compartments. GO analysis showed that half of these genes (16/32) belong to the proteasome family, which plays an important role in both the canonical and non-canonical pathways of NF-κB activation 33. To confirm these findings, we constructed a single cell proteasome score based on the gene expression of proteasome gene set and observed that the proteasome scores in WT glomeruli and cortex were significantly higher than that in the 4KO pigs (figure 4D).
TLR4 and CD40 are essential mediators of NF-κB activation 34, 35, which were upregulated in HB-perfused RECs and significantly reduced in the 4KO RECs (Fig. 3E). To measure if NF-κB regulated genes were also affected, we measured the expression of VCAM by immunostaining and confirmed higher VCAM expression in the HB perfusion WT RECs but less in the 4KO (Fig. 4E). Collectively, our results showed that HB perfusion triggers endothelial activation of genes involved of the NF-κB pathway in WT kidney, while genetic elimination of the porcine MHC class I (SLA-I) and three xenoantigens (GGTA1 CMAH, B4GALNT2) significantly diminishes the activation of these genes (Fig. 4F).
{kind=link}
{kind=link}