Radiation induces widespread transcriptional changes
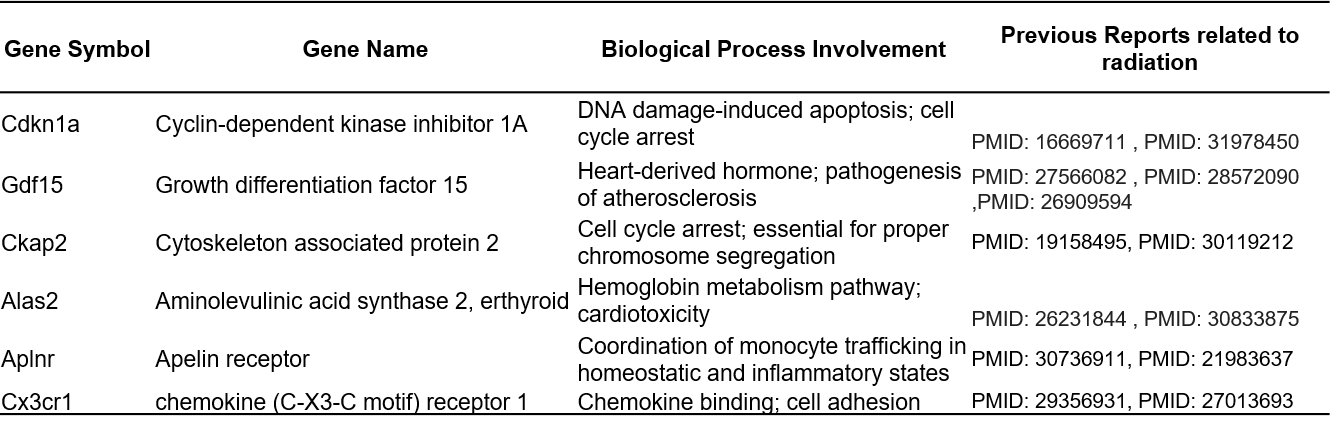
Microarray analysis performed on all mouse heart samples revealed 2041 differentially expressed genes (|log2FC| > 1; p-value < 0.05) that distinguished unirradiated control samples from samples of at least one dose of TBI mice. Overall, most genes had relatively low to no basal expression in control samples and showed increased expression levels after TBI; however, a cluster of genes showed relatively pronounced high expression in control samples that decreased to low expression after irradiation (Fig. 1A). For each dose, more genes were differentially upregulated than downregulated (Fig. 1B). Across all doses, 99 genes were commonly expressed in response to radiation and 128, 55, 390, 322, and 316 genes were expressed exclusively after 1, 2, 4, 8, and 12 Gy of TBI, respectively (Fig. 1C). Supplemental Table S1 lists fold changes and p-values of all differentially expressed genes by dose. While there was no systemic dose-response in terms of the number of genes expressed, we did observe more differentially expressed genes in the higher doses (4, 8, 12 Gy) than in the lower doses (1, 2 Gy). When dose-response of each gene was analyzed by fitting a linear model to each probe, 596 probes were found to have significant dose-responsive up- or down-regulation across all doses (Supplemental Table S2). Cdkn1a, Ckap2, and Gdf15 were among the top 20 probes with the strongest upward linear trend, and Alas2, Aplnr, and Cx3cr1 were among the top 20 probes with the strongest downward linear trend (Fig. 1D). All six genes have previously been reported in the context of radiation or DNA damage response and fall into three main biological roles: cell cycle arrest, hemoglobin metabolism, and inflammatory response (Table 1).
Low basal expression levels of long non-coding RNAs in heart tissue showed increased expression after TBI
In order to understand the response of heart-based lncRNAs to TBI, we filtered whole genome microarray data to include only probes that correspond to transcripts of lncRNAs. Of the 87 lncRNA transcripts in the microarray data that passed the background intensity cutoff in at least one condition, 46 were differentially expressed in response to radiation, irrespective of the TBI dose (Fig. 2A). Most lncRNAs showed relatively low expression in unirradiated control samples with increased expression after radiation. More probes showed upregulation than downregulation in all doses except 2 Gy, which had 4 downregulated lncRNAs and 3 upregulated lncRNAs (Fig. 2A, B). Two lncRNAs were significantly altered at all doses after radiation, systematic names are: chr10:69819062–69871640_F and chr17:29183003–29217681_R (Trp53cor1-up) (Supplemental Table S3) (Fig. 2C). Additional lncRNAs were altered only at specific doses, with 1, 1, 7, 7, and 10 lncRNAs expressed exclusively in 1, 2, 4, 8, and 12 Gy, respectively (Fig. 2C). Supplemental Table S3 lists the fold changes and p-values for the differentially expressed lncRNAs at each dose. Twenty probes showed significant linear upward or downward trends as the dose of TBI increased, demonstrating a linear dose response (Supplemental Table S4). Abhd11os, Trp53cor1, Pvt1, and Kalrn were among the most significant annotated lncRNAs that became upregulated as radiation dose increased, while the linc-RAM (Malrn) transcript had the most significant dose-responsive downregulation (Fig. 2D). Trp53cor1 was the most sensitive to radiation, showing significant increase in the relative intensity in comparison to the unirradiated control even after 1 Gy of TBI. The basal level expression of Trp53cor1 lncRNA expression was below detection threshold levels in unirradiated heart tissue. Due to this reason, we used relative intensity to describe expression of Trp53cor1 after radiation.
In contrast, basal expression of Abhd11os was at a much higher threshold across all doses, including control samples, with significantly higher expression levels after 8 and 12 Gy TBI. Pvt1 showed significance after 2 Gy and Abhd11os showed significance after 8 Gy, while Kalrn and linc-RAM showed significance only after 12 Gy TBI. We confirmed the expression of Dino, Pvt1, and Trp53cor1 in heart samples through RT-qPCR (Fig. 2E). In concordance with the microarray data, Trp53cor1 showed very low expression in control samples but significant dose-responsive upregulation after radiation. Pvt1 also showed consistent results with the microarray in terms of the dose response; however, it showed significance in expression change only after 12 Gy TBI. Damage induced noncoding lncRNA (Dino) was not present in our microarray data due to lack of a probe, but prior data from our lab led us to validate its expression in the heart via RT-qPCR. We found that Dino is also significantly expressed in the heart after every dose of TBI. Like Trp53cor1, Dino showed very low expression levels in control samples but increased significantly after radiation (Fig. 2E).
Low dose of whole-body irradiation induces the most significant and pronounced changes of microRNA expression in mouse heart tissue
A separate whole genome microarray analysis of miRNA expression revealed 102 significantly altered miRNAs in mouse heart tissue in response to radiation. Surprisingly, the largest and most significant changes in expression occurred after 1 Gy of TBI, with 86 differentially expressed miRNAs identified at this dose (Fig. 3A). Furthermore, there were no commonly expressed miRNAs across all doses and no miRNAs significantly expressed in 4 or 8 Gy TBI samples (Fig. 3B, C). We did observe significant regulation of miRNAs at 2 and 12 Gy; however, the numbers were relatively low, with 1 and 19 miRNAs in 2 and 12 Gy, respectively. Fold changes and p-values for all significantly altered miRNAs at each dose are listed in Supplemental Table S5. Linear trends were fit to all miRNA probes to identify dose response across all doses and found 21 probes that showed significant upward linear trend (Supplemental Table S6). No miRNAs showed a significant downward linear trend. Among the top linear probes were miR-149-3p, miR-6538, miR-3960, miR-8101, miR-7118-5p, and miR-211-3p, all of which showed a statistically significant increase in expression only after 12 Gy of TBI (Fig. 3D).
Pathway analysis of mRNA targets reveals biological role of inversely correlated mRNA-miRNA pairs and their potential as part of an integrated TBI response signature
Since we are ultimately interested in developing integrated signatures of coding and non-coding RNA response to radiation, we sought to identify potential interactions of the miRNA and mRNA signatures and their biological significance. Using IPA, we first conducted miRNA target filter analysis of the significant differentially expressed miRNAs to identify experimentally verified mRNA targets in our dataset. Canonical pathway analysis of the identified targets revealed significant activation of pathways relevant to cell cycle checkpoint activation and senescence, including p53 signaling and numerous apoptosis signaling pathways, among others (Fig. 4A). Interestingly, most pathways activated across all doses were predicted to have the highest activation after 4 Gy TBI, followed by lower activation in 8 and 12 Gy. One exception to this was the senescence pathway, which showed the lowest activation at 4 Gy. A concurrent higher activation of various apoptosis pathways after 4 Gy TBI may suggest that miRNA-mRNA pairs at this dose exhibit a different response to stress and DNA damage than both lower and higher doses at this time-point after irradiation. After pathway analysis, we identified predicted miRNA-mRNA pairs with inverse expression patterns, demonstrating their potential for inclusion in an integrated RNA marker signature to improve clinical decision making. We found three miRNAs that showed significant expression in at least one dose and had predicted targets with inverse expression patterns, each miRNA with two targets. Radiation decreased expression of both miR-128-3p and miR-122-5p relative to control, while their targets—Tgfbr1 and Wee1, and Fam117b and Slc7a11, respectively—showed a statistically significant increase in expression as radiation increased (Fig. 4B). The third miRNA, miR-18-5p, showed increased expression after radiation, most significantly after 1 Gy. Its targets, E2f1 and E2f2, showed significant down regulation across all doses.
Pathway analysis of all mRNAs predicts activation of immune and cell-cycle related pathways and deactivation of metabolic pathways after TBI
While understanding the interactions and biological implications of the miRNAs and mRNAs is critical for developing an integrated biomarker signature, we hypothesized that because the relatively low number of differentially expressed miRNAs would limit the number of mRNAs included in the pathway analysis, we could potentially miss genes that play a significant role in pathway regulation. Therefore, we also conducted a canonical pathway analysis using all differentially expressed mRNAs, irrespective of interactions with miRNAs in our dataset. A similar overall pattern of pathway regulation exists between the target mRNAs and target/non-target mRNAs, with significant deactivation of most of the pathways involved (Fig. 5A). Several pathways related to coagulation, including both ex- and intrinsic prothrombin activation and the coagulation system pathway, were downregulated across all doses. Changes in immune-related pathways were less consistent in terms of activation or deactivation. While natural killer cell signaling was activated significantly after 4 and 12 Gy TBI, the complement system was inhibited across all doses.
Two clusters of pathways showed the most pronounced activation and deactivation. Five pathways were predicted to be activated across all doses, and all were involved in immune response or cell cycle regulation. There were 78 genes differentially expressed in these pathways, a majority of which were downregulated with respect to the control (Fig. 5B). A full list of genes can be found in Supplemental Table 7. In contrast to the pathway analysis of the mRNA targets, which showed the highest activation of several pathways after 4 Gy TBI, all activated pathways except natural killer cell signaling were consistently activated as the dose increased. We observed that radiation inhibits pathways relevant to xenobiotic metabolism and biosynthesis of lipids, including hormones. This is demonstrated by activation of LPS/IL-1 mediated inhibition of RXR function and inhibition of the super-pathway of melatonin degradation, among others. There were 79 genes involved in deactivation of these seven pathways, most of which were downregulated after TBI (Fig. 5C, Supplemental Table S7).
{kind=link}
{kind=link}