Screening of efficient mCherry-binding protein (MBP) pairs to design Cre recombinase dependent on RFPs
First, we aimed to construct Cre recombinase dependent on RFPs based on the reported tool named Cre-DOG (Tang et al., 2015). In this system, N-terminal and C-terminal split Cre recombinase fragments are fused with specific nanobodies for target proteins, and target proteins mediate reunion of the split Cre recombinase fragments (Figure 1A). In order to identify functional pairs of binding proteins, we selected 6 nanobodies and 2 DARPins previously reported to have a highly specific binding property to a monomeric RFP, mCherry (Brauchle et al., 2014; Fridy et al., 2014), and we constructed every combination (8 + 8 = 16 constructs). The codon-optimized DNA sequences were synthesized and were inserted instead of GFP-specific nanobodies in pAAV-EF1α-N-CretrcintG (Addgene ID: 69570) or pAAV-EF1α-C-CreintG (Addgene ID: 69571) using NheI and EcoRI sites. We renamed them as MBPs (mCherry-binding proteins) 1 – 8 in this study. MBPs 1-6 are nanobodies and MBPs 7 and 8 are DARPins. As a consequence, we obtained pAAV-EF1α-N-Cre-MBP(1-8)-WPRE and pAAV-EF1α-C-Cre-MBP(1-8)-WPRE.
Then we performed in vitro luciferase reporter assays to find adequate MBP pairs that could induce reunion to reconstruct an active Cre recombinase. N-Cre-MBP, C-Cre-MBP, FLEX-NanoLuc, and target RFPs were co-transfected into HEK293 cells by the calcium phosphate method. The FLEX switch consists of paired loxP and lox2272 sequences and enables the expression of a gene of interest only when Cre recombinase is functional (Atasoy et al., 2008). NanoLuc is a small and bright luciferase from the deep-sea shrimp Oplophorus gracilirostris (Hall et al., 2012). Recombinase activities were measured as luminescence derived from the bioluminescent reaction catalyzed by NanoLuc luciferase. We tested mCherry, mRFP1, and tdTomato as target proteins. These red fluorescent proteins were all derived from the same wild-type DsRed protein (Shaner et al., 2004). While mCherry and mRFP1 are monomeric, tdTomato is a tandem dimer of two subunits. We also tested mRuby, which is a monomeric variant of the red fluorescent protein eqFP611 derived from Entacmaea quadricolor (Kredel et al., 2009), as a negative control. As shown in Figure 1B and 1C, heat maps of mCherry and mRFP1 showed similar patterns. The pair of N-Cre-MBP6 and C-Cre-MBP1 induced high activity for both mCherry and mRFP1. Twin pairs of the same MBP such as N-Cre-MBP1 and C-Cre-MBP1 or N-Cre-MBP2 and C-Cre-MBP2 showed weak reporter activities, possibly indicating competition for the same binding site by both N-Cre-MBP and C-Cre-MBP. On the other hand, the heat map for tdTomato greatly differed from those of mCherry and mRFP1 (Figure 1D). Unfortunately, we did not find any strong signal when we targeted the tetrameric RFP, DsRed (Figure 1E). The twin pair of N-Cre-MBP8 and C-Cre-MBP8 showed the highest activity for tdTomato. The heat maps for mRuby and No RFPs (Figure 1F and 1G) showed only weak recombinase activities around the maps. According to these heat maps obtained from luciferase assays, we selected the pair of N-Cre-MBP6 and C-Cre-MBP1 as a candidate pair for Cre-dependent on monomeric RFP.
Characterization of recombinase activity of Cre-DORs in vitro
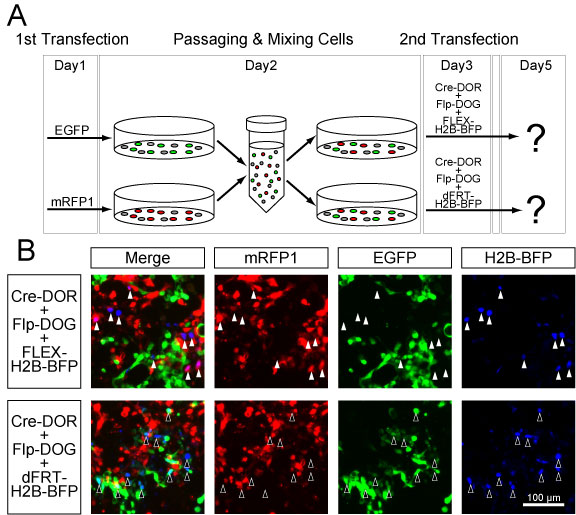
Next, we investigated the recombination efficiency of the pair of N-Cre-MBP6 and C-Cre-MBP1 (Cre-DORN6C1) using a fluorescent protein reporter. Four kinds of plasmids including N-Cre-MBP6, C-Cre-MBP1, target RFPs, and FLEX-H2B-GFP were co-transfected into HEK293 cells by the calcium phosphate method (Figure 2A). H2B-GFP shows nuclear localization because H2B (histone 2B) protein binds to the DNA in the nucleus. Recombinase activities were measured as H2B-GFP expression induced by FLEX switching (Figure 2B). The fluorescent signal of GFP was enhanced by immunostaining using a GFP antibody. Quantitative cell counting of fluorescent images showed that 81.8 ± 1.5% of mCherry-positive cells were GFP-positive and that 74.1 ± 1.6% of mRFP1-positive cells were GFP-positive, while 5.6 ± 0.6% of mRuby-positive cells were GFP-positive (n = 8 each) (Figure 2C, 2D). The recombination efficiency of Cre-DORN6C1 + mCherry was 14.6-times higher than that of Cre-DORN6C1 + mRuby. The Cre-DORN6C1 system was found to be dependent on all three components of target RFPs, N-Cre-MBP6, and C-Cre-MBP1 in the system. Removal of N-Cre-MBP6 or C-Cre-MBP1 resulted in total loss of reporter activity. Cell counting showed that 0.9 ± 0.1% of mRFP1-positive cells were GFP-positive without CCre-MBP1 and that 0.8 ± 0.1% of mRFP1-positive cells were GFP-positive without NCre-MBP8 (n = 8 each). In all cases, the percentages of RFP-positive cells in GFP-positive cells were higher than 90% possibly because of the transfection method (mCherry: 98.9 ± 0.3%, mRFP1: 98.3 ± 0.3%, mRuby: 91.0 ± 1.9%, mRFP1ΔCCre: 94.4 ± 3.7%, mRFP1ΔNCre: 88.5 ± 5.6%).
We also tested the recombination efficiency of the pair of N-Cre-MBP5 and C-Cre-MBP4 (Cre-DORN5C4). Quantitative cell counting of fluorescent images showed that 57.9 ± 1.6% of mCherry-positive cells were GFP-positive and that 9.3 ± 1.1% of mRFP1-positive cells were GFP-positive, while only 0.9 ± 0.1% of mRuby-positive cells were GFP-positive (n = 8 each) (Figure 2E, 2F). The recombination efficiency of Cre-DORN5C4 + mCherry was 61.0 times-higher than that of Cre-DORN5C4 + mRuby. The Cre-DORN5C4 system was also found to be dependent on all three components of target RFPs, N-Cre-MBP5, and C-Cre-MBP4 in the system. Removal of N-Cre-MBP5 or C-Cre-MBP4 resulted in total loss of reporter activity. Cell counting showed that 0.7 ± 0.1% of mRFP1-positive cells were GFP-positive without CCre-MBP4 and that 0.8 ± 0.2% of mRFP1-positive cells were GFP-positive without NCre-MBP5 (n = 8 each). Although Cre-DORN5C4 can induce more specific recombination dependent on mCherry, we decided to use Cre-DORN6C1 in the later in vivo experiments because of its high efficiency when it was applied for mRFP1.
In the heatmap of the luciferase assay shown in Figure 1D, we found that the pair of N-Cre-MBP8 and C-Cre-MBP8 shows the highest recombination efficiency dependent on tdTomato. Therefore, we also tested the recombination efficiency of the pair of N-Cre-MBP8 and C-Cre-MBP8 (Cre-DORN8C8). Quantitative cell counting of fluorescent images showed that 46.9 ± 1.1% of tdtomato-positive cells were GFP-positive, while 12.8 ± 1.2% of mRuby-positive cells were GFP-positive (n = 4 each) (Figure 2G). Although Cre-DORN8C8 shows mild specificity against tdTomato, we thought that the S/N ratio of Cre-DORN8C8 was not sufficiently high for use in vivo.
Cellular Localization Of Target Rfps Affects Cre-dor Activity
Cre recombinase exerts its activity within the nucleus. Therefore, it is possible that cellular localization of target RFPs affects Cre-DOR recombinase activity. In order to test this hypothesis, we created mCherry and tdTomato fused with various localization signal peptides: CAAX motif for membrane localization, NES for cytosolic localization, and NLS for nuclear localization (Figure 3A). We observed clear intracellular translocalization of mRFP1 and tdTomato by being fused with these motifs. mCherry/tdTomato fused with CAAX were localized in the plasma membrane, mCherry/tdTomato fused with NES were localized in the cytosol, mCherry/tdTomato without any motif were localized in both the cytosol and nucleus, and nls-mCherry/tdTomato were localized in the nucleus (Figure 3B). Next, we performed in vitro luciferase reporter assays to investigate the effect of cellular translocation on recombinase activity. Plasmids for N-Cre-MBP, C-Cre-MBP, FLEX-NanoLuc, and target RFPs were co-transfected into HEK293 cells. When Cre-DORN6C1 or Cre-DORN8C8 targets RFPs with localization signals, the luciferase assay showed a clear difference between the cellular localizations (Figure 3C). A gradual incremental tendency of recombination activity among membrane-bound, cytosolic, and nucleic localization of target RFPs was observed. Quantitative analyses of luciferase assay data showed that recombinase activity is significantly different between Cre-DORN6C1/Cre-DORN8C8 + mCherry/tdTomato-NES and Cre-DORN6C1/Cre-DORN8C8 + nls-mCherry/tdTomato (P < 0.0001, Tukey’s multiple comparison test, n = 11). Note that average fluorescence intensity of mCherry-NES or tdTomato-NES is higher than that of nls-mCherry or nls-tdTomato.
Next, we aimed to control Cre-DOR activity by using chemical ligands. Glucocorticoid receptor (GR) is a nuclear receptor and it is translocated into the nucleus after binding its ligand, glucocorticoid (Lupien et al., 2009) (Figure 3D). Ligand-induced translocation of GR has been detected by addition of a GFP to the N-terminus of GR (Htun et al., 1996; Mikuni et al., 2007). Therefore, we attached RFPs to human glucocorticoid receptor alpha (hGRα) in order to control Cre-DOR activity. We found that mCherry-GR was localized in the cytosol without its ligand, dexamethasone (Dex) and that it translocated to the nucleus after incubation with Dex (1 µM, 1 hour) (Figure 3E). In Figure 3E, nuclei are visualized by fluorescence from Histone 2B-BFP (pAAV-EF1α-H2B-BFP-WPRE) that was co-transfected with mCherry-GR. Cre recombinase activity of Cre-DORN6C1 targeting mCherry-GR was strongly increased by the presence of Dex (Figure 3F).
An increase in Cre recombinase activity by nuclear localization of RFPs suggests that Cre-DOR activity can be manipulated by controlling the intracellular localization of target RFPs. In order to test this hypothesis, we used a light-inducible nuclear export domain called LEXY (Niopek et al., 2016). LEXY consists of an engineered LOV2 domain from Avena sativa phototropin-1 (AsLOV2), in which the C-terminal Jα helix was converted into an artificial NES. In the dark, the NES is tightly packed against the AsLOV2 core and is thus inactive. Exposure to blue light induces unfolding of the modified Jα helix, uncovering the NES (Figure 3G). We confirmed that nls-mCherry-LEXY was localized mainly in the nucleus in the dark, and blue light illumination (465 nm, 7 W/m2) induced translocation of fused RFPs into the cytosol in 30 min (Figure 3H). In Figure 3H, nuclei are visualized by fluorescence from Histone 2B-BFP that was co-transfected with nls-mCherry-LEXY. We found that blue light illumination inhibited Cre recombination activity of Cre-DORN6C1 targeting nls-mCherry-LEXY as indexed by luciferase activity (Figure 3I).
Functional assay of Cre-DOR in vivo using AAV vectors
In order to examine whether Cre-DORN6C1 functions in living animals, we generated AAV vectors encoding N-Cre-MBP6, C-Cre-MBP1, FLEX-nlsGFP, and target RFPs. GFP tagged with a nuclear localization signal (nlsGFP) is localized mainly in the nucleus. 600 nl of a mixture of virus vectors for Cre-DORN6C1, FLEX-nlsGFP (1 x 1012 vg/ml each), and target mRFP1 (5 x 1010 vg/ml) was unilaterally injected into the right side M1 cortex of wild-type 10-week-old male mice (Figure 4A). The titer of the target RFP-expressing vector was lowered to induce scattered expression and make it easy to count fluorescent protein-expressing cells separately. Recombinase activities were measured as nlsGFP expression induced by FLEX switching (Figure 4B). Four weeks after injection, mice were sacrificed for immunohistochemistry and brain slices were stained with anti-GFP. Clear expression of nlsGFP at the injected site in the M1 cortex was observed (Figure 4C). Quantitative cell counting of fluorescent images showed that 50.2 ± 2.5% of mRFP1-positive cells were GFP-positive and that 93.5 ± 0.6% of GFP-positive cells were mRFP1-positive (n = 5 each) (Figure 4D, 4E). The expression efficiency of GFP in the center area of injection was higher than that in the peripheral area. In order to check the specificity of the Cre-DORN6C1 system in vivo, we injected vectors for Cre-DORN6C1, FLEX-nlsGFP (1 x 1012 vg/ml each), and mRuby (5 x 1010 vg/ml) as a control (Figure 4F). While we observed comparative amounts of mRuby-expressing neurons in the M1 cortex, we found only sparce nlsGFP-expressing neurons in the same area (Figure 4G). Quantitative cell counting of nlsGFP and mRuby-positive cells showed a clear difference between Cre-DORN6C1 + mRFP1-injected mice and Cre-DORN6C1 + mRuby-injected mice. The cell counting showed that 1.5 ± 0.5% of mRuby-positive cells were GFP-positive and that 13.3 ± 4.1% of GFP-positive cells were mRuby-positive (n = 5 each) (Figure 4D, 4E). The recombination efficiency of Cre-DORN6C1 + mRFP1 was 34.4-times higher than that of Cre-DORN6C1 + mRuby in these experiments. All these data suggest in vivo specificity of the Cre-DORN6C1 system using AAV vectors.
Functional Assay Of Cre-dor In Mrfp1-expressing Transgenic Mice
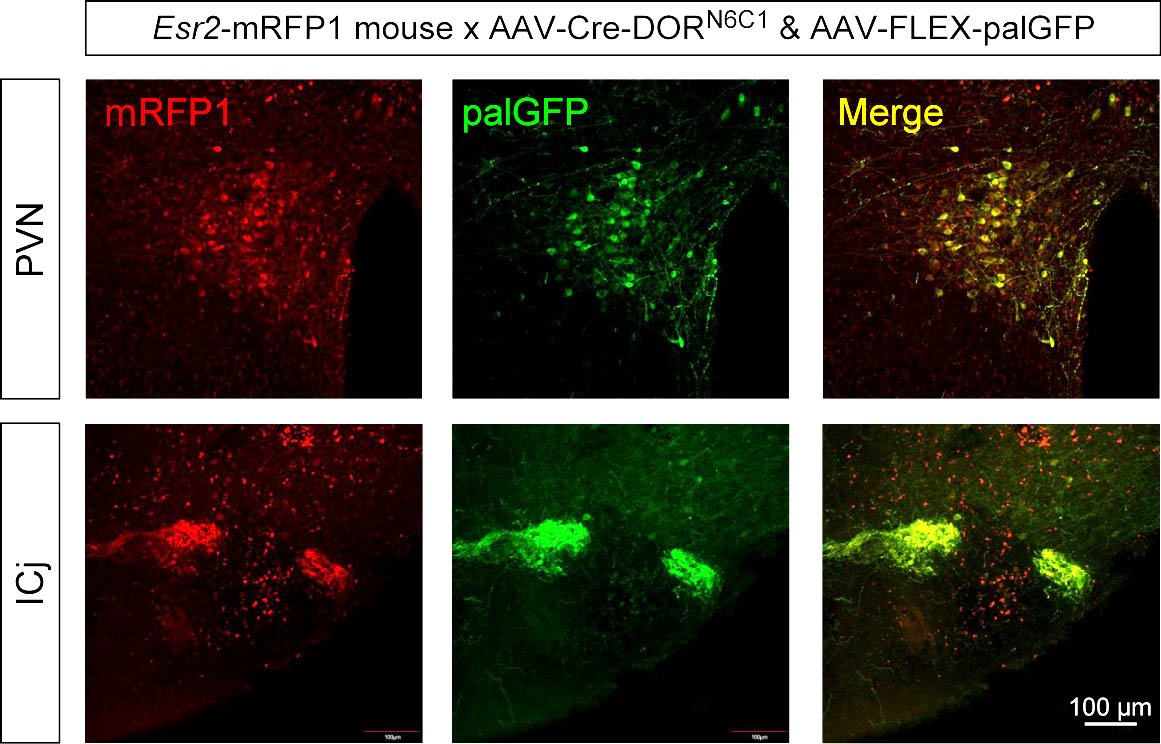
Next, we examined selective expression by Cre-DORN6C1 in mRFP1-expressing transgenic animals. In Esr2-mRFP1 mice, neurons in the paraventricular nucleus (PVN) are visualized by mRFP1. 1 µl of a mixture of AAV9-EF1α-NCre-MBP6-WPRE (6 x 1012 vg/ml), AAV9-EF1α-CCre-MBP1-WPRE (6 x 1012 vg/ml) and AAV9-CAG-FLEX-palGFP-WPRE (6 x 1012 vg/ml) was injected in the PVN of Esr2-mRFP1 transgenic mice (Figure 5A). Recombinase activities were measured as expression of GFP tagged with a palmitoylation signal (palGFP) induced by FLEX switching (Figure 5B). palGFP is sorted to the plasma membrane and has been used to trace neuronal fibers anterogradely (Furuta et al., 2001; Nasanbuyan et al., 2018). Four weeks after injection, mice were sacrificed for immunohistochemistry and brain slices were stained with anti-GFP and anti-mRFP1. Clear expression of palGFP at the injected site in the PVN was observed (Figure 5C). It has been reported that mRFP1-expressing neurons in the PVN of Esr2-mRFP1 mice include oxytocin neurons (Sagoshi et al., 2020) and oxytocin neurons send their axons into the posterior pituitary. In accordance with these previous findings, we observed clear projection from the palGFP-expressing neurons in the PVN and dense axonal terminals in the posterior pituitary (Figure 5D). Quantitative cell counting of palGFP-positive cells in the PVN showed a clear difference between mRFP(+) mice and mRFP(-) mice (Figure 5E). The cell counting showed that 24.1% of mRFP1-positive neurons in the PVN express palGFP on average (Figure 5F). These results showed the usability of Cre-DORN6C1 for detection of selective neural projection in mRFP1-expressing transgenic animals. We also confirmed that Cre-DORN6C1 can be functional in other parts of the brain such as the islands of Calleja (ICj) in Esr2-mRFP1 mice (Supplemental Figure 1).
Anterograde tracing of mRFP1-expressing neurons in Grpr-mRFP1 rats
Finally, we examined selective expression by Cre-DORN6C1 in mRFP1-expressing transgenic animals. In gastrin-releasing peptide receptor (Grpr)-mRFP1 transgenic rats, neurons in the posterior amygdala are visualized by mRFP1. 1 µl of a mixture of AAV9-EF1α-NCre-MBP6-WPRE (6 x 1012 vg/ml), AAV9-EF1α-CCre-MBP1-WPRE (6 x 1012 vg/ml) and AAV9-CAG-FLEX-palGFP-WPRE (6 x 1012 vg/ml) was injected in the medial amygdala area of male Grpr-mRFP1 transgenic rats (Figure 6A). Four weeks after injection, rats were sacrificed for immunohistochemistry and brain slices were stained with anti-GFP and anti-mRFP1. Clear and selective expression of palGFP at the injected site in the posterodorsal medial amygdala (MePD) was observed (Figure 6B). We observed clear projection from the palGFP-expressing neurons in the MePD. We found a bundle of smooth passing fibers in the stria terminalis (ste) and dense axonal terminals with varicosity in the posterior bed nucleus of the stria terminalis (BSTp or STMP) (Figure 6C). These findings suggest that mRFP-expressing neurons in the MePD send their axons to the BSTp (Figure 6E). Finally, we confirmed this neural projection using a retrograde tracer. We injected 300 nl of green retrobeads in the BSTp of Grpr-mRFP1 transgenic rats (Figure 6D). One week after injection, rats were sacrificed. We detected some mRFP1 neurons that included green retrobeads in the MePD. These results support our idea that mRFP-expressing neurons in the MePD send their axons to the BSTp (Figure 6F).
{kind=link}
{kind=link}