3.1 MTT assays
The effect of CQDs/Cu2O, Cu2O on cell cytotoxicity was tested in cancer cells (HeLa, A549, HT-29, SKOV3, HCT116), and normal cells (BABL-3T3, HEK293T, J774A1) by the MTT assay. Cells were treated with CQDs/Cu2O or Cu2O (1.56, 3.12, 6.25, 12.5, 25 µg mL-1) for 24 -72 h. The effect of CQDs was detected in HeLa, SKOV3 and BABL-3T3 cells by MTT assay. The results were shown CQDs were non-toxic and biocompatibility, which is agreed with previous findings. 34 As shown in Figure 1, CQDs/Cu2O displayed cytotoxicity in a concentration-dependent manner in all tested cells, and SKOV3 cells displayed the highest inhibitory rate for proliferation among all cells. To further verify the results of MTT assay, a second mitochondrial activity-based assay, WST-1, was employed to measure the cytotoxicity of CQDs/ Cu2O in SKOV3 cells. WST data showed that CQDs/Cu2O inhibited the growth of SKOV3 cells in a dose-dependent manner, the value of IC50 obtained by WST assay (IC50 = 1.46 µg mL-1) was consistent with the result from MTT assay (IC50 = 1.50 µg mL-1). It was suggested the MTT assay can be used to evaluate the cytotoxicity of CQDs/Cu2O.
Figure 1. Differential cytotoxicity of CQDs/Cu2O in cancer cells (HeLa, A549, HT-29, SKOV3, HCT116) and normal cells (BABL-3T3, HEK293T, J774A1) by the MTT assay for 24 h.
Interestingly, when the concentration of CQDs/Cu2O was lower than 12.5 µg mL-1, a discriminative difference was identified for the cytotoxicity between SKOV3 cancer cells and other cancer cells, even normal BABL-3T3, HEK293T and J774A1 cells. This is important because developing an anticancer drug that can effectively distinguish between tumor and normal cells is currently the greatest challenge.21
In general, the CQDs/Cu2O composite displayed cytotoxic activity against cancer and normal cells in the IC50 (half maximal inhibitory concentration) range of 0.85 to 22.4 µg mL-1 (Table 1). The IC50 value of CQDs/Cu2O against cells was lower than Cu2O and CQDs were non-toxic. It was indicated that the cytotoxicity of the CQDs/Cu2O composite was higher than that of the CQDs and Cu2O. In addition, the leakage of copper into the medium was undetectable at 37 °C after 72 h by ICP-AES (coupled plasma-atomic emission spectrometry) measurement, suggesting CQDs/Cu2O displayed high stability and the CQDs/Cu2O plays a leading roles in cytotoxic activity. Moreover, the IC50 value of CQDs/Cu2O against SKOV3 cells was lower than above mentioned cancer cells and normal cells, which was approximately 3-fold lower than other tested cancer cells and approximately 12-fold lower than normal cells. It was further suggested CQDs/Cu2O was sensitive to SKOV3 cells.
Table 1. Comparison of the IC50 (μg mL−1) of CQDs/Cu2O composite and Cu2O in different cell lines.
|
Cells
|
IC50 (CQDs/Cu2O)
|
IC50 (Cu2O)
|
|
24 h
|
48 h
|
72 h
|
24 h
|
48 h
|
72 h
|
|
HeLa
|
10.6
|
9.5
|
4.9
|
27.8
|
23.9
|
28.1
|
|
A549
|
15.8
|
12.3
|
10.5
|
31.3
|
24.7
|
23.6
|
|
HT-29
|
7.1
|
4.0
|
2.5
|
16.2
|
14.3
|
15.5
|
|
SKOV3
|
1.5
|
0.9
|
0.85
|
15.7
|
13.9
|
12.2
|
|
HCT116
|
9.4
|
6.9
|
5.8
|
15.0
|
11.7
|
14.8
|
|
BABL-3T3
|
18.9
|
17.8
|
16.9
|
29.5
|
27.4
|
18.3
|
|
HEK293T
|
22.4
|
16.3
|
11.7
|
24.8
|
19.6
|
15.9
|
|
J774A1
|
20.1
|
16.4
|
14.2
|
23.8
|
18.9
|
16.6
|
On the other hand, compared with commercial anticancer agents such as ART and OXA at different times, the cytotoxicity of CQDs/Cu2O was better than that of ART and OXA against cancer cells (Table 2). The IC50 of CQDs/Cu2O in SKOV3 cells (IC50= 0.85 µg mL -1) was approximately 75-fold lower than that of OXA (IC50= 64.6 µg mL -1) 21 and 114-fold lower than that of ART (IC50= 96.9 µg mL -1) 21 at 72 h. Therefore, CQDs/Cu2O may have a potential utility for effectively distinguishing between SKOV3 cells and other tested cells.
Table 2. Comparison of the IC50 (μg mL−1) of CQDs/Cu2O, OXA and ART in different cell lines.
|
Cells
|
IC50 (CQDs/Cu2O)
|
IC50 (OXA)
|
IC50 (ART)
|
|
24 h
|
48 h
|
72 h
|
24 h
|
48 h
|
72 h
|
24 h
|
48 h
|
72 h
|
|
HeLa
|
10.6
|
9.5
|
4.9
|
84.821
|
28.521
|
20.021
|
126.521
|
62.721
|
43.621
|
|
A549
|
15.8
|
12.3
|
10.5
|
219.821
|
128.321
|
27.021
|
160.721
|
66.021
|
31.121
|
|
HT-29
|
7.1
|
4.0
|
2.5
|
47.81
|
32.65
|
26.82
|
63.41
|
50.24
|
43.60
|
|
SKOV3
|
1.5
|
0.9
|
0.85
|
241.521
|
120.821
|
64.621
|
280.821
|
121.121
|
96.921
|
|
HCT116
|
9.4
|
6.9
|
5.8
|
62.9
|
41.2
|
28.6
|
40.3
|
22.6
|
21.4
|
|
BABL-3T3
|
18.9
|
17.8
|
16.9
|
77.821
|
34.221
|
13.821
|
118.221
|
52.421
|
36.621
|
3.2 RNA‑Seq data and identification of differentially expressed genes (DEGs)
Six sequencing libraries including CQDs/Cu2O treatment libraries (CQDs1, CQDs2, CQDs3) and control group libraries (Control1, Control2, Control3) were successfully constructed and subsequently sequenced (Table 3). The RNA-Seq of six samples produced 37.95 million raw reads and 37.05 million high quality clean reads. It was shown that 95.47-97.77% of the reads were successfully mapped to the Homo genome (Table 3). Compared to the control, fragments per kilobase of transcript per million fragments (FPKM) analysis indicated that a total of 1251 transcripts at a fold change >2 and false discovery rate (FDR) <0.05 were differentially expressed, among which, 495 genes were upregulated while 756 were downregulated (Figure 2).
Table 3. The raw data from RNA-Seq analysis of the CQDs/Cu2O and control group.
|
Sample
|
Raw reads
|
Clean reads
|
Total mapped
|
Q30 (%)
|
GC content (%)
|
|
Control1
|
56611380
|
54934068
|
52445514(95.47%)
|
93.71
|
50.23
|
|
Control2
|
68457650
|
66554960
|
63613738(95.58%)
|
93.97
|
50.25
|
|
Control3
|
60574334
|
59402794
|
58070632(97.76%)
|
96.09
|
50.49
|
|
CQDs1
|
63142992
|
61528818
|
58951428(95.81%)
|
94.14
|
50.40
|
|
CQDs2
|
65707806
|
63946010
|
61244702(95.78%)
|
93.97
|
50.23
|
|
CQDs3
|
65006850
|
63690950
|
62268500(97.77%)
|
96.08
|
50.88
|
In order to determine the differentially expressed genes at lower doses and elucidate the mechanism of selective inhibition of SKOV3 cells by CQDs/Cu2O, the RNA sequencing analysis were performed after SKOV3 cells treated with 3.12 μg mL-1 CQDs/Cu2O for 24 h. Compared to the control, fragments per kilobase of transcript per million fragments (FPKM) analysis indicated that a total of 1011 transcripts at a fold change >2 and false discovery rate (FDR) <0.05 were differentially expressed, among which, 381 genes were upregulated while 630 were downregulated. There was no significant difference between the high concentration group and the low concentration group. On the one hand, the enriched GO terms in the DEGs of cells treated by 3.12 μg mL-1 CQDs/Cu2O have also been provided in supporting information (Figure S1). As shown in Figure S1, the distribution of enriched GO terms of cells treated by 3.12 μg mL-1 CQDs/Cu2O was similar with that treated by 12.5 μg mL-1 CQDs/Cu2O. Moreover, DEGs analysis results showed that the angiogenesis-related genes (Maspin and TSP1) were significantly upregulated after 3.12 μg mL-1 CQDs/Cu2O treatment, which was also consistent with those of higher doses treatment.
Figure 2. Volcano plot of DEGs in SKOV3 cells after treated with CQDs/Cu2O. The red dots expressed the upregulated genes and green dots expressed the downregulated genes. The black dots indicate the genes with no significant differential expression.
3.3 GO and KEGG analysis of DEGs
Gene Ontology (GO) was used to analyze the obtained DEGs, where three main ontologies such as biological process, cellular component, and molecular function performed. As shown in Figure 3, the top 20 most enriched GO terms are summarized. In the category of biological process, the most abundant groups were “cellular process”, “single-organism process”, and “biological regulation”. Within the cellular component category, the “cell”, “cell part”, and “organelle” were identified as the most enriched GO terms. The molecular functional groups of DEGs were also related to “binding”.
Figure 3. The enriched GO terms in the DEGs of cells treated by 12.5 μg mL-1 CQDs/Cu2O. The green, red, and blue bars represent the terms of biological process, cellular component, and molecular function, respectively.
GO enrichment analysis was also used to identify significantly enriched BP, CC, or MF terms in DEGs during CQDs/Cu2O treatment. As shown in Table 4, there were 10, 8, and 6 ontology terms enriched in BP, CC, and MF, respectively. The enriched BP terms mainly included the regulation of the apoptotic process, programmed cell death, and cell motility, indicating that CQDs/Cu2O plays a key role in the induction of SKOV3 cell apoptosis and the regulation of cell movement. The enriched CC terms mainly included proteinaceous extracellular matrix and integral components of the plasma membrane, suggesting that CQDs/Cu2O affected the extracellular matrix and plasma membrane. The enriched MF terms included calcium ion binding, nucleic acid binding, receptor binding, etc.
Table 4. GO enrichment analysis of the DEGs during CQDs/Cu2O treatment (P value<0.01)
|
Description
|
GO ID
|
Term Type
|
Number
|
P-value uncorrected
|
P-value corrected
|
|
Regulation of apoptotic process
|
GO:0042981
|
BP
|
138
|
1.89891E-10
|
0
|
|
Regulation of programmed cell death
|
GO:0043067
|
138
|
2.3484E-10
|
0
|
|
Regulation of locomotion
|
GO:0040012
|
91
|
2.44938E-10
|
0
|
|
Regulation of cell death
|
GO:0010941
|
150
|
2.78134E-10
|
0
|
|
Regulation of localization
|
GO:0032879
|
216
|
2.82001E-10
|
0
|
|
Response to organic substance
|
GO:0010033
|
168
|
3.03004E-10
|
0
|
|
Positive regulation of response to stimulus
|
GO:0048584
|
184
|
3.05792E-10
|
0
|
|
Regulation of cell motility
|
GO:2000145
|
84
|
3.254E-10
|
0
|
|
Positive regulation of signal transduction
|
GO:0009967
|
134
|
3.27693E-10
|
0
|
|
Anatomical structure development
|
GO:0048856
|
293
|
1.89891E-10
|
0
|
|
Proteinaceous extracellular matrix
|
GO:0005578
|
CC
|
57
|
5.76E-11
|
0
|
|
Extracellular matrix
|
GO:0031012
|
70
|
1.49E-10
|
0
|
|
Integral component of plasma membrane
|
GO:0005887
|
129
|
1.9E-10
|
0
|
|
Plasma membrane part
|
GO:0044459
|
227
|
2.99E-10
|
0
|
|
Intrinsic component of plasma membrane
|
GO:0031226
|
136
|
3.4E-10
|
0
|
|
Extracellular space
|
GO:0005615
|
153
|
3.46E-10
|
0
|
|
Extracellular region
|
GO:0005576
|
219
|
3.7E-10
|
0
|
|
Extracellular region part
|
GO:0044421
|
321
|
3.82E-10
|
0
|
|
Calcium ion binding
|
GO:0005509
|
MF
|
79
|
6.87E-10
|
0
|
|
Nucleic acid binding
|
GO:0003676
|
145
|
1.02E-09
|
0
|
|
Heparin binding
|
GO:0008201
|
26
|
1.12E-07
|
0
|
|
Receptor binding
|
GO:0005102
|
127
|
1.67E-07
|
0
|
|
Sulfur compound binding
|
GO:1901681
|
32
|
2.52E-07
|
0
|
|
Glycosaminoglycan binding
|
GO:0005539
|
29
|
9.63E-07
|
0.002
|
To determine the effect of CQDs/Cu2O on the functions of DEGs, the KEGG data base was used to analyze annotated pathways of DEGs. The result indicated that DEGs were significantly enriched in 20 pathways after treated with CQDs/Cu2O in Figure 5 (P corrected <0.05). According to the number of DEGs, the three pathways such as steroid biosynthesis, focal adhesion, and ECM-receptor interaction were the most enriched pathways. Based on the lowest P value, steroid biosynthesis was among the most notable enriched pathways.
Figure 4. A bubble diagram of the significant enrichment pathways through KEGG analysis of DEGs (P < 0.05). The size and color of bubble indicated the number of gene enriched in certain pathway and the P-value, respectively. The rich factor = (the number of DEGs mapped to a certain pathway)/ (the total number of genes mapped to this pathway).
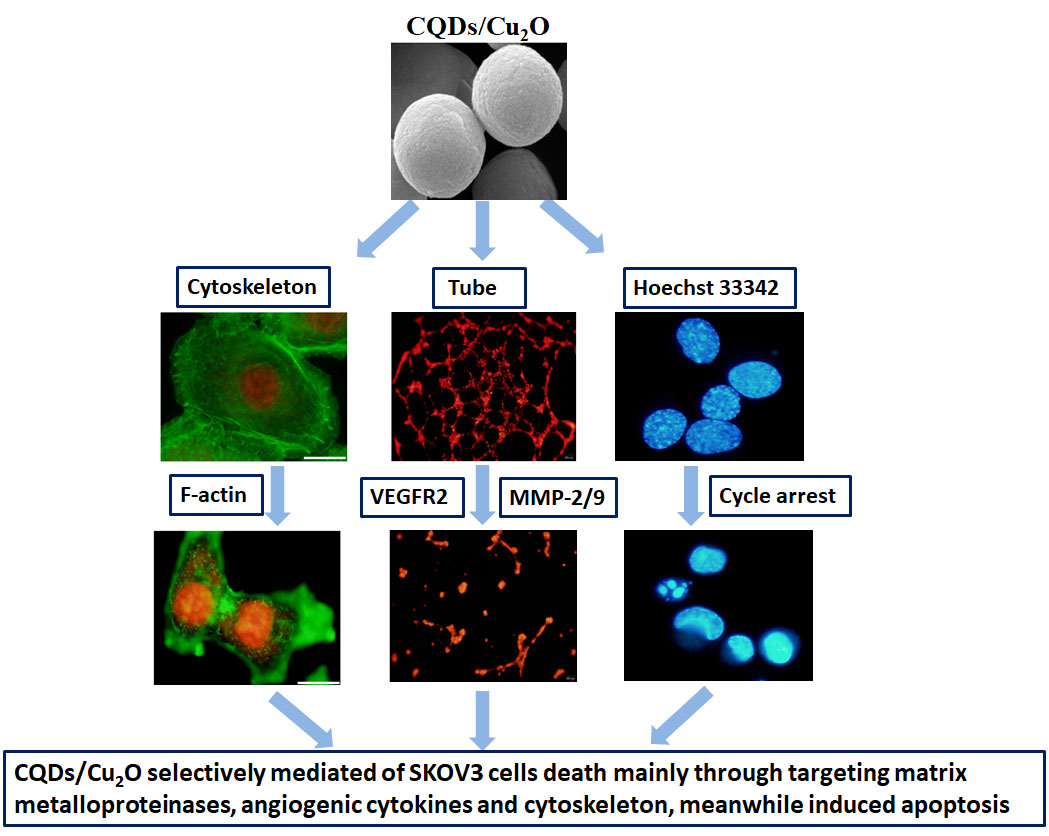
3.4 CQDs/Cu2O induced apoptosis of SKOV3 cells
Transcriptome analysis revealed that apoptotic process and programmed cell death were significantly affected. AO/EB (acridine orange/ethidium bromide), Hoechst 33342, and FITC-Annexin-V/PI were used to demonstrate apoptosis. AO is absorbed by viable and nonviable cells to emit green fluorescence, while EB is absorbed by nonviable cells to emit red fluorescence. Therefore, viable cells displayed uniform bright green fluorescence with organized structure. The apoptotic cells showed red to orange fluorescence with fragmented chromatin, and the necrotic cells showed red fluorescence with swollen. 44 As shown in Figure 5, cells appeared healthy with a green nucleus in the control group. Some of the cells incubated with the CQDs/Cu2O samples had an orange nuclei and fragmented chromatin, indicating that they were either in necrosis or at the late stage of apoptosis. In addition, only few necrotic cells with uniformly red nuclei and an organized structure appeared after treatment with the same concentration of Cu2O. However, there was no significant change in the CQD group. These results indicate that CQDs/Cu2O induces necrosis or apoptosis of SKOV3 cells, and the effect of the CQDs/Cu2O composite is better than that of CQDs and Cu2O.
Figure 5. Fluorescence photomicrographs changes by AO/EB staining of SKOV3 cells after treatment with CQDs/Cu2O, CQDs, or Cu2O for 24 h compared to controls, respectively.
The apoptotic effects of CQDs/Cu2O on SKOV3 cells were determined by staining with Hoechst 33342. Nuclear condensation & DNA fragmentation are considered to be typical features of apoptosis. SKOV3 cells were treated with 6.25 μg mL−1 CQDs/Cu2O for 24 h. As shown in Figure 6, cells displayed a uniformly blue fluorescence in the control group and CQDs group. CQDs/Cu2O treated cells showed nuclear shrinkage, cytoplasmic blebbing, and chromatin condensation, including bright blue dots in the nuclei, thereby representing nuclear fragmentation, whereas a small number of cells were chromatin condensation in Cu2O group. These observations further indicated CQDs/Cu2O can induce SKOV3 cells apoptosis, which is better than both CQDs and Cu2O.
Figure 6. Fluorescence photomicrographs changes by Hoechst 33342 staining of SKOV3 cells after treatment with CQDs (B), Cu2O (C) and CQDs/Cu2O (D) for 24 h compared to controls (A), respectively. Scale bar 20 μm.
A flow cytometric analysis via FITC-Annexin-V/PI assay was used to further confirm the CQDs/Cu2O-induced apoptosis. As shown in Figure 7A, as the concentration of CQDs/Cu2O increased from 3.12~25 μg mL−1, the proportion of cells in the upper right quadrants increased from 3.5% to 48.4%, however, the proportions of other quadrant cells did not change significantly. This data also confirm that CQDs/Cu2O can effectively induce SKOV3 cell apoptosis.
Figure 7. Effects of CQDs/Cu2O on the apoptosis and cell cycle distribution by flow cytometry. (A) Flow cytometric analysis of SKOV3 cells incubated with CQDs/Cu2O for 24 h using an apoptosis kit with dual fluorescence of annexin V-FITC/PI. (B) SKOV3 were treated with CQDs/Cu2O for 24 h and stained using PI to determine the content of DNA. The experiments were tested at least 3 times.
3.5 Determination of the Cell Cycle
The cell cycle, which includes DNA replication, mitosis, and cytokinesis, is a major event in cell division. It has been proven that deregulation of the cell cycle is related to numerous carcinogenic processes.45 Nanomaterials, such as Se, Ag, ZnO, Ag-ZnO, TiO2 NPs, have been reported to arrest cell cycle to suppress cancer cell proliferation.46-49 Cu-based nanomaterials also play a critical role in regulating the cell cycle. For example, cuprous oxide nanoparticles (CONPs) suppressed the proliferation of HeLa cells and caused cell cycle arrest in G0/G1.36 A previous study reported that Cu-MOF (HKUST-1) has the potential to inhibit the proliferation of SKOV3 by cell cycle arrest in G2/M.30 To determine whether apoptosis is involved in the cell cycle effect of CQDs/Cu2O, SKOV3 cells were treated by CQDs/Cu2O and subjected to flow cytometry analysis (Figure 7B). As shown in Figure 7B, CQDs/Cu2O led to accumulation of cells in the S phase (9.24 ± 0.9% in control group versus 23.8 ± 0.5% in 12.5 μg mL-1 CQDs/Cu2O treated group (P < 0.001), while there was no significant change in the G0/G1 and G2/M phase. The results showed that the SKOV3 cells are arrested at S phase after treatment by CQDs/Cu2O, indicating that mitosis and proliferation of cancer cells were inhibited.
3.6 Effects of CQDs/Cu2O on the SKOV3 migration
Ovarian cancer is the one of the leading causes of death in gynecological malignancies. Local recurrence and metastasis are considered to be the main causes of treatment failure for ovarian cancer. It was critical to suppress the metastasis of cancer cells for the development of effective drugs and therapies. GO enrichment analysis revealed that CQDs/Cu2O regulated cell motility. Hence, the wound-healing assay was used to detect the amount of migrated cells and the width of scratch after treatment with CQDs/Cu2O. As shown in Figure 8A, the width of scratch increases with increasing concentrations of CQDs/Cu2O and prolonging reaction time. As well as the amount of migrated cells decreased after treated with CQDs/Cu2O (Figure 8B). The migration inhibition rate increased from 61.8% to 79.4% after incubation with CQDs/Cu2O (Figure 8C). When cells treated with 25 μg mL-1 CQDs/Cu2O, the migration inhibition rate doesn't show significant change at 6, 12 and 24h, mainly due to the strong cytotoxicity of high concentration of CQDS /Cu2O, and the inhibitory effect on cell migration was shown in a short time and maintained for a period of time. CQDs/Cu2O was demonstrated to exhibit remarkable concentration- and time-dependency, suggesting that it has the potential to suppress SKOV3 cell migration.
We found that the expression of MMP-2/9 sharply decreased after treatment with CQDs/Cu2O for 24 h by western blot analysis (Figure 8D). It has been reported the decrease of MMPs expression is considered to be the main factor for the inhibition potential of migration and angiogenesis. Carbon nanocapsules (HMCNs) and fullerene-based nanoparticle Gd@C82(OH)22 serve as potent migration inhibitors by downregulating MMP-2/9.20, 50 The results further indicated that CQDs/Cu2O effectively suppressed SKOV3 cell migration by regulating the expression of metalloproteinase.
Figure 8. Effect of CQDs/Cu2O on SKOV3 cell migration by the wound-healing assay. (A) The “scratch” was produced by scraping the monolayer SKOV3 cells using a pipette tip after grown to form a confluent monolayer. (Scale bar 100 μm). The relative migration activity (B) and the inhibition rate of migration (C). The experiments were tested at least 3 times.
3.7 Effects of CQDs/Cu2O on HUVEC blood vessel formation
Angiogenesis is required for tumor progression and metastasis to provide oxygen and nutrients. Targeting angiogenesis has become a unique perspective and strategy in anticancer therapy. DEGs analysis results showed that the SERPINB5 (known as Maspin) and THBS1 (known as TSP1) genes were significantly upregulated after CQDs/Cu2O treatment. To confirm whether CQDs/Cu2O has an anti-angiogenesis effect, a blood vessel formation in vitro model system was selected and the effect of CQDs/Cu2O on new blood vessel formation in HUVECs cells was assessed. VEGF was used to induce endothelial cell proliferation, migration, and differentiation into blood vessel structures. As shown in Figure 9A, a robust and complete blood vessel produced after HUVECs cells treated with VEGF within 12 h. CQDs/Cu2O effectively inhibited the length of blood vessel. A reduction of approximately 40~70% in total blood vessel length per field following treatment with 3.12 to 12.5 μg mL−1 CQDs/Cu2O for 6 h. When the time was extended to 12 h, the inhibition percentage increased from 60 to 90%, suggesting a dose-dependent and a time-dependent decrease. The viability of HUVEC cells was also determined by MTT assay. It was shown that the inhibition rate was lower than 20% when HUVEC cells were treated with CQDs/Cu2O for 12 h (Figure S2), indicating that this concentration of CQDs/Cu2O was non-toxic over a short period and antiangiogenic activity was not caused by the cytotoxicity of CQDs/Cu2O.
Moreover, semaxanib (SU5416, 87 μM), as a selective inhibitor of VEGF, was used as the experimental control. CQDs/Cu2O also had better antiangiogenic activity than SU5416 and Cu2O (Figure 9B). In Figure 9C, the length of blood vessel induced by CQDs/Cu2O (12.5 μg mL−1) was about 1.8-fold shorter than the length of blood vessel induced by SU5416. The length was approximately 4.2-fold shorter than that caused by Cu2O. CQDs/Cu2O inhibited tube formation by ~90%, which was more than that with Cu2O (53%) and SU5416 (80%) (Figure 9D). It has been reported 80 μg mL−1 AgNPs19 suppressed about 80% blood vessel formation of bovine retinal endothelial cells. 12.5 μg mL−1 Fe-MIL-101 21 effectively suppressed about 60% blood vessel formation of HUVEC cells. It was suggested that CQDs/Cu2O possessed better anti-angiogenic activity than AgNPs and Fe-MIL-101.
A variety of growth factors and cytokines are involved in regulating angiogenesis. VEGF, as one of the most important pro-angiogenic factors, combined to receptors VEGFR1 and VEGFR2, stimulating endothelial cell migration and blood vessel formation.13, 17, 51 As shown in Figure 8D, the expression of VEGFR2 decreased after treatment with CQDs/Cu2O. Altogether, our results indicated that angiogenesis mediated by VEGFR2 and MMP-2/9, indicating that CQDs/Cu2O may be a potential candidate in anti-angiogenic therapy.
Figure 9. Dil-labeled HUVECs were inoculated in Matrigel-coated wells and induced blood vessel formation. Scale bar, 200 μm. (A) CQDs/Cu2O composite inhibited blood vessel formation. Scale bar, 200 μm. (B) The effect of 12.5 μg mL-1 CQDs/Cu2O (87 μM), Cu2O (87 μM) and SU5416 (87 μM) on blood vessel formation. (C) Cumulative tube length in four fields/well and the inhibition percentage of the tube length.
3.8 CQDs/Cu2O induced alterations in the cytoskeleton of SKOV3 cells
The cytoskeleton, which has a variety of biopolymer networks such as F-actin, microtubules, and intermediate filaments, plays a fundamental role in all eukaryotic cells. 52 Dynamic regulation of the F-actin cytoskeleton is essential for many physiological cellular processes, including cell adhesion, migration, division, and apoptosis of cancer cells. We investigated the effect of CQDs/Cu2O on the F-actin cytoskeleton of SKOV3 cells. First, we observed the morphological changes of the F-actin cytoskeleton by FITC-conjugated phalloidin and PI immunofluorescence staining. As shown in Figure 10, SKOV3 cells in the control group displayed normal actin structure, single intact nucleus, and short F-actin bundles arranged around the cell membrane. After treated by CQDs/Cu2O, a large number of fluorescence spots were scattered throughout the cytoplasm, gradually disappeared as the CQDs/Cu2O concentration increased and the fluorescence intensity decreased, which significantly differed from that in normal F-actins. CQDs/Cu2O might therefore exhibit enhanced damage and disruption of F-actin. We also evaluated the content of F-actin cytoskeleton after treated by CQDs/Cu2O. The results of western blot demonstrates that the expression of F-actin in SKOV3 cells treated with CQDs/Cu2O significantly was significantly lower than that in the control group (Figure 8D). It has been reported that some nanoparticles exhibit depolymerization effects or disrupt the cytoskeletal architecture and affect cell division, such as GO nanosheets, 24 ZnO nanoparticles,25 gold nanoparticles, 53 silver nanoparticles,54 and MOFs (IRMOF-3).28 Compared to the above nanoparticles and agents, we could demonstrate the disruption of the cytoskeleton filament owing to CQDs/Cu2O exposure and prove that the expression of the F-actin cytoskeleton decreased at the protein level. Hence, the suppression of SKOV3 migration by CQDs/Cu2O may be related to the downregulation of F-actin. It may be unique perspective and strategy in anticancer therapy.
Figure 10. Effects of CQDs/Cu2O-stimulated disruption of the cytoskeleton. SKOV3 cells were stained using FITC-conjugated phalloidin and PI after treatment with CQDs/Cu2O (0, 6.25, 12.5, 25 μg mL-1) for 24 h. Images are representative of at least three experiments, Scale bar, 20 μm.
{kind=link}