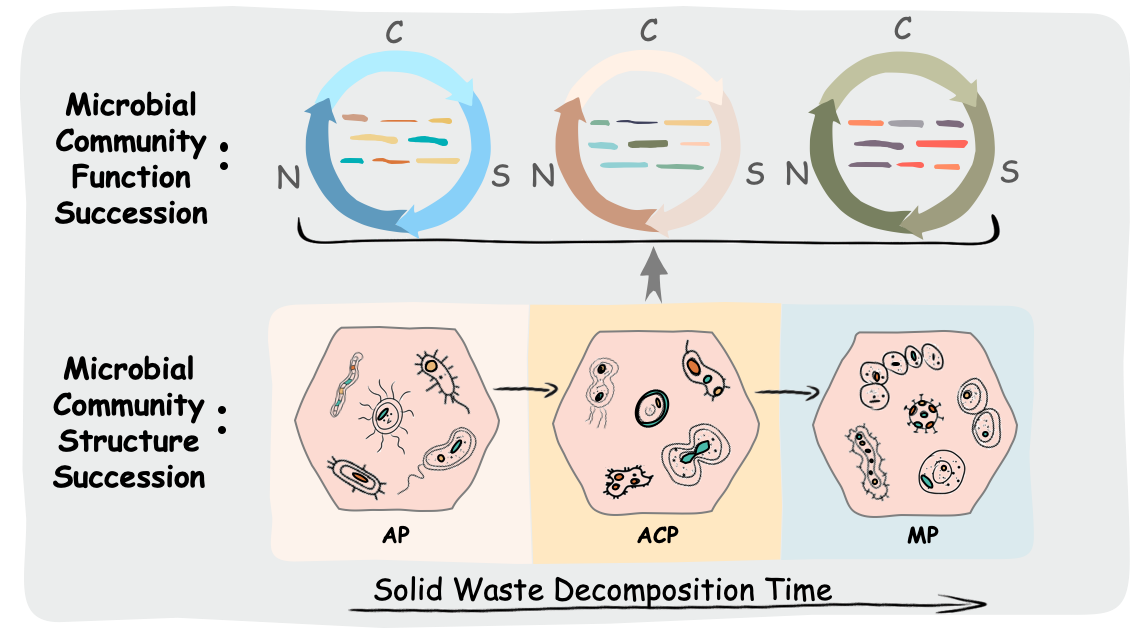
Landfill, a “baby” terrestrial ecosystems, represents a significant carbon sink with carbon biotransformation [7]. Meanwhile, landfill releases large amount of microorganism-mediated greenhouse gas (CO2, CH4, and N2O). Therefore, landfill is a good example to study the biological mechanisms that regulate carbon exchanges between the belowground and atmosphere. In this study, integration of solid waste physiochemical parameters, 16S rRNA based prosequencing and metagenomics analysis, we show that the structure and potential function of microbial communities changed along solid waste decomposition in response to biodegradable substrates.
Liable carbon seems to maintain long term carbon stability and storage (Zhou et al., 2012). During solid waste decomposition, microorganism first hydrolyze cellulose and food waste to from hydrolysis products (sugars, amino acid and fatty acid), then metabolize hydrolysis products to methane metabolic precursors (acetate and H2), and finally convert methane metabolic precursors to methane [7]. Accordingly, microbial community composition changed according to the available biodegradable substrate. For instance, the relative abundance of Firmicutes assigned by 16S rRNA pyrosequencing data set, typical cellulose-degrading microorganisms [36], increased from 32.5 ± 2.0% in AP to 45 ± 3.2% in ACP and then decreased to 13.2 ± 1.3% in MP, suggesting its important roles in hydrolysis and fermentation. The relative abundance of Firmicutes assigned by metagenomic data set was also lower in MP (11.1%) than AP (35%) and ACP (31.1%). Both metagenomic and 16S rRNA prosequencing data sets showed that Methanosarcinales and Methanomicrobiales were main contributors in methanogenesis to produce methane and enrich in MP. Our previous study [37] showed that BOD5 significantly influenced bacterial and archaeal communities structure in solid waste during solid waste decomposition. Because hydrolysis, the rate-limited step on solid waste decomposition, mainly contributes to the levels of BOD5, it can be concluded that biodegradable substrate is key factor in shaping microbial community structure during solid waste decomposition.
Carbon cycling during solid waste decomposition can be seen from the variation in related genes involved in various carbon cycling pathways. As expected, genes (L-lactate dehydrogenase) involved in fermentation enriched in the AP and ACP, whereas genes (mcrB and methyl coenzyme M reductase system, component A2) involved in methanogensis occurred in ACP and became abundant in MP. This variation accords to the changes of available biodegradable substrate as well as microbial community composition during solid waste decomposition. Anaerobic C fixation (related genes 2-oxoglutarate: ferredoxin oxidoreductase subunit alpha and beta, frdA, ATP citrate lyase, CO dehydrogenase subunit delta and gamma) was the main pathway in carbon fixation in solid waste than aerobic C fixation (RubisCo small chain and phosphoribulkinase) owing to the anoxic condition in solid waste bioreactor. Substrate quality impact on microbial assimilation of soil organic carbon that higher substrate quality leads to higher ratio of microbial carbon to soil organic carbon [2]. In this study, comparison of the loss of OM and BDM% indicated that recalcitrant carbon was stored in solid waste. OM includes liable and recalcitrant carbon, while BDM% represents biodegradable carbon. There are 56-63% biodegradable carbon (indicated by BDM%) lost, much higher than13-28% liable and recalcitrant carbon lost (indicated by OM).
Greenhouse gas (CO2, CH4, and N2O) emission from solid waste can be reflected by their related genes variation during solid waste decomposition. Methane emission mainly occurred in MP with abundant genes (mcrB and methyl coenzyme M reductase system, component A2) involved in methanogensis. During this period, genes methane monooxygenase involved in aerobic CH4 oxidation occurred by 100% Corynebacteriales. Carbon dioxide emission by aerobic respiration (coxI, coxIII, coxA, and coxC) occurred across solid waste decomposition and showed an increasing trend. In AP, oxygen in void space of solid waste was consumed rapidly. In ACP and MP, aerobic respiration became strong. This is because oxygen carried by recirculation leachate was consumed, which is consistent with the status of in situ leachate recirculation in landfill that leachate recirculation improved carbon dioxide production [38]. N2O emission by denitrification occurred across solid waste decomposition. The involved genes (nosB, nosC, and norZ) increased across this period, leading to the increasing nitrate lost. Oxygen diffusion into solid waste by leachate recirculation is supposed to inhibit denitrification. However, denitrification activity became strong along solid waste decomposition. This is probably because strong aerobic respiration consumed diffused oxygen rapidly, making aerobic condition transiently existed.
N availability affects the carbon sink activity of terrestrial ecosystems. Microbial N2 fixation increased under elevated CO2 [39, 40]. A previous functional gene-based GeoChip analysis showed that the abundance of nifH genes for N2 fixation increased under elevated CO2 but net nitrification, net N mineralization, and the total soil N content were not significantly changed [39]. In this study, N2 fixation were detected during solid waste decomposition and the relative abundance of genes (nifD, nifH, nifK, and nitrogenase) decreased along solid waste decomposition, from 2.7% in AP, to 1.7% in ACP, and to 1.2% to MP, suggesting dynamics of N2 fixation. As we did not measure the changes of CO2, it is difficult to relate the genes to the changes of CO2. The link between genes of N2 fixation and system-level process requires further study. Nitrification were not detected in this study, which is agree with the high ammonia accumulation in the bioreactor. This can partly explained the “ammonia accumulation” in landfill or leachate [38, 41]. Anammox was also not detected in this study. Although anammox is assumed to exist in bioreactor landfill [42] and Anammox bacterium Candidatus Kuenenia stuttgartiensis [43] was found in aged solid waste, there is absent of functional genes evidence.
Sulfur also affects the carbon sink activity of terrestrial ecosystems [1, 44]. A previous large-scale inter-region analysis of time series data showed that dissolve organic carbon (DOC) was controlled by the changes of sulfate [44]. High potential for assimilatory sulfate reduction (related genes: cysC, cysD, and cysN) and sulfur mineralization (related genes: cysteine dioxygenase and 3-mercaptopyruvate sulfurtransferase) by Rhodobacterales, Pseudomonadales, Propionibacteriales, and Bacteroidales was detected across solid waste decomposition. These two pathways converted sulfate to organic sulfur and to hydrogen sulfide. Dissimilatory sulfate reduction and sulfide oxidation by Clostridiales directly convert sulfate to hydrogen sulfide, although the abundance of their genes (aprA, aprB, and dsrA) was low in this study. Polysulfide is thought to be a possible intermediate that used by bacteria in sulfur respiration [45]. Bacterium Clostridium were able to reduce polysulfide to hydrogen sulfide and also produced acetate when lactate was supplied [46]. In this study, the gene (psrA) for polysulfide reduction was abundant in ACP, suggesting more hydrogen sulfide released in this phase. Taken together, these sulfur cycle pathways co-contribute hydrogen sulfide release in landfill. The relationship between sulfate and DOC in leachate should be investigated in the future.
Like bacteria, archaea are now recognized to have important roles in global C and N cycles [47]. In this study, the potential for C cycling (methanogensis, anaerobic carbon fixation and Calvin cycle) and N cycling (nitrogen assimilation, mineralization and N2 fixation) involved by archaea were extensively detected. Autotrophic archaea are either anaerobes or can tolerate or use oxygen only at low concentrations [48]. They can oxidize inorganic substrates (e.g., H2, H2S, CO, NH3 etc) and use inorganic compounds (e.g., SO42− and NO3−) as electron acceptors [48]. Those inorganic substrates and compounds enriched in landfill where is also anoxic condition, providing habitat for autotrophic archaea. Autotrophic carbon fixation is via different metabolic pathways. Key gene phosphoribulkinase and RubisCo for Calvin cycle was present in Methanomicrobiales in this study as well as others [49]. However, none of the chemolithoautotrophic archaea seems to use this cycle for CO2 fixation [48]. Genomes of Methanosaeta thermophile contains genes encoding phosphoribulkinase and RubisCo (genomes available from the DOE joint Genome Institute website) [49], suggesting potential ability of Calvin cycle. Key genes such as 2-oxoglutaratesythase and ATP-citrate lyase involved in reductive citric acid cycle and CO dehydrogenase for Wood-Ljungdahl pathway [48] were also present in Methanosarcinales in MP and Methanosarcinales and Methanomicrobiales in ACP in this study, suggesting autotrophic archaea paly important role in anaerobic carbon fixation under anoxic condition. Archaean also involved in N2 fixation, including Methanosarcinales [50] and Methanomicrobiales [47]. There is a shift between bacteria and archaea involved in N2 fixation across solid waste decomposition. Bacteria (Clostridiales and Bacteroidales) was initial dominant in AP and ACP. Archaea (Methanosarcinales) abundance increased along solid waste decomposition and became dominant in methanogenic phase, reflecting the role of archaea in N2 fixation became important in methanogensis.
Potential C, N, and S cycling pathways during solid waste decomposition coupled with bacterial and archaea groups. Specific, Clostridiales were detected as main contributors in several pathways. Clostridiales, capable of decomposing cellulose, have been extensively detected in landfill and leachate [51–53]. However, their role in C and N cycling has not been elucidated. In this study, Clostridiales has been assigned as potential catalyzing anaerobic carbon fixation, fermentation, aerobic CO oxidation, nitrogen assimilation, mineralization, and ammonification, N2 fixation, dissimilatory sulfate reduction, and assimilatory sulfate reduction, suggesting Clostridiales play important role in C, N, and S cycling in landfill.
For long term perspective, carbon cycle in landfill requires consideration of complex interactions and feedbacks that occur between microbes, plants (natural or anthropogenic plantation application after landfill closed) and their physical environment in the context of climate change. Up to date, carbon dioxide and methane flux from landfill have been studied, whereas carbon storage in landfill and N2O emission received little attention. In particular, the mechanism of microbial mediation of carbon cycling are poorly understood. Given the expanding of landfills, it is urgent to understand the mechanisms by which microorganisms regulate the carbon exchange between landfill-atmosphere.
In conclusion, the issues addressed in this study are important to the understanding of carbon dynamics in landfill. We found that bacterial and archaeal community structure and functional genes involved in C and N fixation and greenhouse gas emission changed during solid waste decomposition. Such shifts in bacterial and archaeal community structure and function could potentially modify the direction and magnitude of landfill regulation of the rate of carbon storage and greenhouse gas emission, thereby influence the climate change. Our findings also have practical consequence that these observed special bacterial and archaeal groups mediated C, N, and S cycles during solid waste decomposition provided direct microbiological evidence for landfill stabilization, thereby showing clues for carbon and nitrogen management and pollution control in landfill.
{kind=link}