A total of 41.17 Gb of high-quality paired-end reads were obtained by Illumina genomic sequencing (~92.22X coverage, Table S1). The genome size of M. dirhodum was estimated to be 457.2 M based on k-mer counting. The k-mer distribution analysis revealed a peak at 79.8× of the sequencing depth, suggesting a moderate level of heterozygosity (0.445%) and highly repetitive sequence content (59.20%) in the genome (Fig. 1A), which was similar to other Aphidinae insects with low or moderate level heterozygosity11,18. To obtain a reference genome for M. dirhodum, we generated 161.53 Gb of PacBio long reads using CCS model (Table S1), which were subsequently corrected into 10.34 Gb HiFi reads. The genome was initially assembled using hifiasm, resulting in 296 contigs with a contig N50 of 7.82 Mb and the longest contig of 23.64 Mb (Table 1). A total of 41.17 Gb of short reads generated by Illumina NovaSeq 6000 platform were then mapped against our assembly, resulting in a mapping rate of 92.18%. The BUSCO analysis showed that 96.9% (single-copied gene: 92.5%, duplicated gene: 4.4%) of 1,367 single-copy genes in the insecta_odb9 database were identified as complete, 0.4% of genes were fragmented, and 2.7% of genes were missing in the assembled genome. The percentage of complete single-copy genes is higher than those in the genomes of some other insect species, such as S. miscanthi (90.2%)11, R. maidis (94.5%)18, A. pisum (93.5%)16 and E. lanigerum (96.8%)17. Considering the moderate level of heterozygosity and the high level repetitiveness of the genome, the current result represents a high-quality genome assembly of M. dirhodum.
For the chromosome-level assembly, 38.09 Gb of clean reads (150 bp paired-end) were obtained from the Hi-C library (coverage: 85.31X, Table S1). Totally 118,367,396 (86.83%) reads were mapped to the draft genome and 96,331,684 (70.67%) of them were uniquely mapped. The uniquely mapped sequences were analyzed with 3D-DNA software to assist genomic assembly. As a result, 68 scaffolds were assembled with an N50 length of 37.54 Mb (Table 1). Finally, a total of 447.8 Mb genomic sequences (accounting for 98.50% of the whole assembled length) were located on 9 chromosomes (Fig. 1B, Table 1, Table S2), supporting a 2n = 18 karyotype for M. dirhodum, which is identical to S. miscanthi11. The contig N50, Scaffold N50 of M. dirhodum were also much higher than that of previous reported aphid genome assemblies (Table 1). This is the first high-quality chromosome-level genome of M. dirhodum, which will be very helpful for cloning, functional verification and evolutionary analysis of genes in this important species, or even other Hemiptera insects.
Genome Annotation
The repeatmasker and repbase were used to annotate the repeat sequences. In total, 34.97% of the M. dirhodum genome was annotated as repeat sequences. Long terminal repeats (LTRs), long interspersed nuclear elements (LINEs) and DNA transposons accounted for 9.23%, 2.25% and 10.33% of the whole genome, respectively, and 13.16% of repeat sequences were annotated as unclassified. A total of 286 tRNAs were predicted by trnascan-SE. Using infernal, we also identified 51 small nucleolar RNAs (snoRNAs), 586 ribosomal RNAs (rRNAs), 73 small nuclear RNAs (snRNAs), 59 microRNAs (miRNAs), 286 tRNAs and 639 other types of ncRNAs.
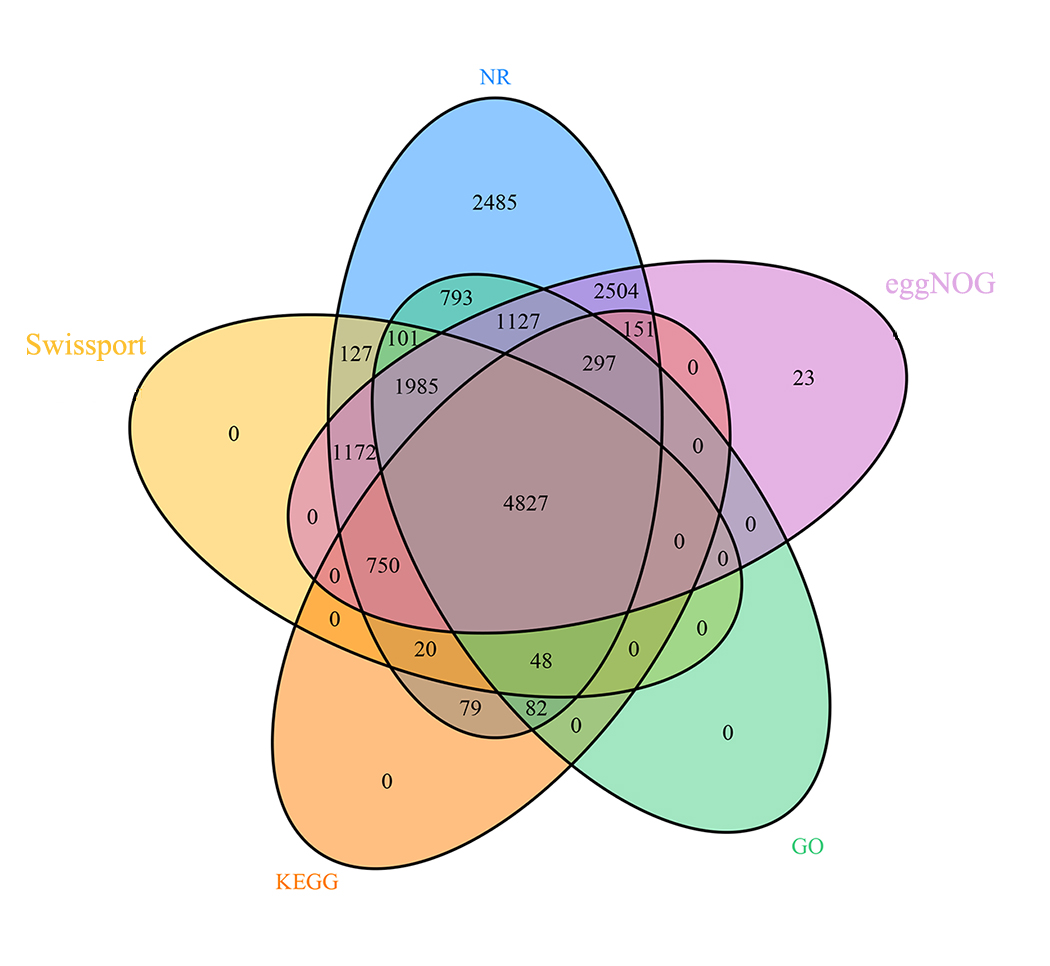
After masking repeat sequences, a total of 18,003 protein-coding genes with the mean CDS length of 1,776 bp were identified from the M. dirhodum genome using de novo, homology- and RNA sequencing-based methods. The number of genes in the M. dirhodum genome is comparable to that of several other Aphidinae species, such as 16,006 protein-coding genes in S. miscanthi11 and 19,097 protein-coding genes in Diuraphis noxia23, but far less than R. maidis18, A. pisum16, M. persicae24 and E. lanigerum17, which has 26,286, 36,195, 23,910 and 28,186 protein-coding genes, respectively (Table 1). Functional annotation found that 16,548 (91.92%), 9,030 (50.16%), and 12,836 (71.30%) genes had significant hits with proteins catalogued in NR, SwissProt and eggNOG, respectively. There were 9,260 (51.44%) and 6,254 (34.74%) genes annotated to GO terms and KEGG pathway, respectively (Fig. S1).
Genome synteny and Phylogeny analysis
To gain insights into an evolutionary perspective for M. dirhodum, a whole genome-based phylogenetic analysis was performed with eight other hemipteran insect species, including M. persicae24, D. noxia23, A. pisum16, R. maidis18, Melanaphis sacchari (GCA_002803265.2), Nilaparvata lugens25, Bemisia tabaci26,27 and Apolygus lucorum28. Drosophila melanogaster29 was used as the outgroup. A total of 209,881 genes to 22,945 orthogroups for the 10 species comparing was assigned (Fig. 2). Phylogenetic tree was constructed using the single-copy orthologous genes (Table S3). As a result, M. dirhodum and the five other Aphididae insects formed an Aphididae cluster, which showed that M. dirhodum is close to A. pisum, and separated from M. sacchari and R. maidis18. Three other Hemiptera insects including B. tabaci26,27, N. lugens25 and A. lucorum28 formed another cluster (Fig. 2).
Syntenic relationships between M. dirhodum and A. pisum genome were compared. The results reveal high levels of genome rearrangement between chromosomes of M. dirhodum and A. pisum, and a number of fission and fusion events were observed16. Chr1 in M. dirhodum shares 81.9% of the syntenic blocks of chr X in A. pisum (Fig. 3). Considering the conservation of X chromosome in Aphidini insects (Biello et al., 2021), we inferred that chr 1 might be the sex chromosome in M. dirhodum. In addition, chrA1 in A. pisum is mainly syntenic to chr2, chr4, chr5 and chr8 of M. dirhodum; ChrA2 in A. pisum is mainly syntenic to chr6, chr7 and chr9 of M. dirhodum; ChrA3 in A. pisum is mainly syntenic to chr3 in M. dirhodum, respectively16. However, many fusion events covering small regions occurred in all chromosomes between these two insect species (Fig. 3).
Ecdysone signaling is involved in wing dimorphism in M. dirhodum
All aphids are born through viviparous reproduction with wing primordia, but it degenerates by the second instar in the unwinged morph30. In the winged individuals, the wing primordia continue to slowly develop from first to third instar nymphs, and then rapidly grow in the fourth instar. In M. dirhodum, the winged and wingless individuals can be distinguished from the third instar nymph to the adult under a microscope31,32.
RNA-seq was performed between winged and wingless M. dirhodum in the third, fourth instar nymphs and adults using our assembled genome as a reference. Based on our transcriptome and qPCR results, several important genes involved in ecdysone signaling were identified to be different expressed between wingless and winged M. dirhodum (Fig. 4; Table S4, S5, S6). In detail, transcripts annotated as ecdysone receptor (ECR), ecdysone-induced protein 74A (E74A), broad-complex core protein (BR-C) and nuclear hormone receptor FTZ-F1 (FTZ-F1) were up-regulated in wingless individuals in third (Fig. 4A; Table S4) and fourth (Fig. 4B; Table S5) instar nymphs, while ECR and FTZ-F1 were up-regulated in wingless adults (Fig. 4C; Table S6), compared to the winged ones. Ecdysone titers between wingless and winged populations from third instar nymph to adult stages were also measured by enzyme immunoassay (EIA). The wingless individuals from third instar nymphs (1.59-fold), fourth instar nymphs (2.01-fold) and adults (1.47-fold) exhibited higher ecdysone levels compared to the winged controls (Fig. 5).
Ecdysone is an important steroid hormone that controls molting and metamorphosis in insects and even arthropods33,34. Recent studies showed that ecdysone signaling may also play a role in controlling wing-morph determination21. For example, application of ecdysone or its analogues would produce fewer winged offspring in A. pisum, while inhibiting ecdysone signaling could produce more winged offspring35. These results revealed that ecdysone signaling is crucial for transgenerational plasticity in the wing-dimorphic aphid A. pisum. In the present research, effect of ecdysone on wing dimorphism within one generation was investigated. High titers of ecdysone were detected in wingless M. dirhodum. Furthermore, the expression levels of its receptor ECR, together with the downstream genes (E74A, BR-C and FTZ-F1) were all up-regulated in wingless M. dirhodum. That is, ecdysone may promote the expression of E74A, BR-C and FTZ-F1 through ECR, and then induce more wingless individuals (Fig. 6). These investigations lay an important foundation for comprehensive analysis and understanding of ecdysone signaling regulation on wing polymorphism.
Other potential genes involved in wing dimorphism in M. dirhodum
Insulin/IGF-1 signaling (IIS) pathway was also thought to be involved in regulating wing polyphenism21,36,37. Two putative insulin receptors (InRs), InR1 and InR2, have been shown to play opposing roles in wing-morph determination by regulating the activity of the forkhead transcription factor, FOXO, in N. lugens. Activation in nymphs of InR1 results in the suppression of FOXO, and the development of the long winged morph in the adult. Activation of the other receptor (InR2) suppresses the activation of InR1, leading to active FOXO and the development of the short winged morph38,39. Two transcripts annotated as insulin receptor (insulin-like receptor and insulin-like peptide receptor) were also identified in M. dirhodum, which is consistent with the annotation of the A. pisum genome16. The comparative transcriptome analysis showed that the transcript annotated as insulin-like receptor (InR2) was up-regulated in third (2.65-fold) and fourth (2.37-fold) instar wingless nymphs (Table S4, S5), compared to the winged individuals. In addition, one transcript annotated as insulin receptor substrate 1 (InR1) was down-regulated in both third (-3.42-fold) and fourth (-2.68-fold) instar wingless nymphs (Table S4, S5). No DEGs associated with insulin signalling pathway was identified between the wingless and winged M. dirhodum adults (Table S6). FOXO was also identified to be up-regulated in wingless M. dirhodum in third (3.25-fold) and fourth (3.34-fold) instar nymphs and adults (4.03-fold) (Table S4, S5, S6). These results suggest that insulin receptor/FOXO may play an important role in the early wing dimorphism of M. dirhodum.
Wnt2, Fng (fringe), Uba1(Ubiquitin-activating enzyme E1), Hh (hedgehog), Dpp (Decapentaplegic), Brk (Brinker), Ap (alar process), Dll (Distal-less), Hth (helix-turn-helix), Tsh (thyroid-stimulating hormone), Nub (nubbin), Scr (Sex combs reduced), Antp (Antennapedia), Ubx (Ultrabithorax), Asc, Srf (serum response factor) and Fl (flugellos) are important in the wing development of insects, such as Drosophila41,42,43,44, Bombyx mori45, M. persicae46 and A. pisum47. These genes were also identified to be different expressed between wingless and winged M. dirhodum by transcriptome sequencing (Table S4, S5, S6), indicating their potential roles in regulating wing dimorphism or development in M. dirhodum. In addition, a great deal of genes related to muscle composition, energy metabolism and reproduction were also identified between the winged and wingless aphids.
Phylogenetic analysis of Cytochrome P450, CarE and GST families in M. dirhodum
A total of fifty Cytochrome P450 (CYP) genes were identified from our assembled genome and the full-length transcriptome sequences. According to the standard nomenclature, all these P450s were divided into 12 families and 22 subfamilies (Fig. 7, Table S7). The largest families included CYP6 with 20 genes and CYP380 with 11 genes (Table S7). Chromosome 8 had the largest P450 cluster with 5 genes (CYP6CY1, CYP6CY4, CYP6CY5, CYP6CY6 and CYP6CY90). In the closely related Aphididae species A. pisum, fifty-eight P450s are found in 12 families and 24 subfamilies16. Phylogenetic analysis among M. dirhodum and two other Aphididae species (A. pisum and R. maidis) resolved the four expected P450 groups including the CYP2, CYP3, CYP4 and mitochondrial clans (Fig. 7). Only CYP6 family was identified in the CYP3 clan in M. dirhodum, including 6 subfamilies (CYP6CY, CYP6CZ, CYP6DC, CYP6DD, CYP6DB and CYP6YC) (Fig. 7, Table S7). Most P450s in CYP6 family showed multiple expansions with close phylogenetic relationships, particularly in the clusters containing CYP6CY (Fig. 7). Previous research has shown that CYP6CY subfamily confer resistance to nicotine in M. persicae48, indicating their important role in host adaptation. In M. dirhodum CYP4 clan, 2 families (CYP4 and CYP380) and 7 subfamilies were identified (Fig. 7, Table S7). Members of the CYP4 clan showed wide diversity in their sequences and exhibited multiple expansions with close phylogenetic relationships. The CYP2 clan contained fewer genes and less evolutionary differentiation than CYP3 and CYP4 clans (Fig. 7). Members of the CYP2 clan are essential for the basic physiological functions in insects. For example, CYP18A1 is tightly correlated with ecdysteroid synthesis49; CYP303A1 is indispensable for embryonic development and adult eclosion50. The mitochondrion clan is generally small and four P450s in 4 families of CYP301, CYP302, CYP315 and CYP353 were found in M. dirhodum (Fig. 7, Table S7). Previous studies have proposed that mitochondrial P450s have two types of functions in insects. One is essential for physiological functions, such as the Halloween genes CYP302A1, and CYP315A1 that produce the cyclic ecdysteroid pulses triggering moulting; The other is rapidly evolved taxon-specific paralogous P45051.
Twenty carboxylesterase (CarE) genes were identified from our assembled genome. All these CarEs were divided into 6 classes (Fig. 8), including two acetylcholinesterases (AChEs), one neuroligin (Nlg), one gliotactin (Gli), three α-esterases (αEs), eight juvenile hormone esterases (JHEs) and five β-esterases (βEs). A closely related duplicate of JHEs (JHE1-JHE8) in M. dirhodum was identified, which is similar to A. pisum16. JHE is a critical enzyme that helps to regulate JH levels, and thus affect the normal development and metamorphosis of insects52.
Twenty-two glutathione S-transferase (GST) genes were obtained from our assembled genome. All these GSTs were divided into 5 classes (Delta, Omega, Sigma, Theta and Microsomal) (Fig. 9). GSTs, especially the insect-specific delta and epsilon classes, play a key role in the metabolism of xenobiotics53. Generally, these two classes of GSTs have largely expanded in insects. However, no member of Epsilon class was identified from M. dirhodum genome, and other Aphididae insect genomes. 9 Delta GSTs were identified in the current M. dirhodum genome, and 8 of them are closely clustered on Chromosome 9, indicating their potentially important roles in M. dirhodum.
Detoxification genes differentially expressed between winged and wingless M. dirhodum
For the transcriptome results, 11 P450s, 12 CarEs, 7 GSTs, 15 UDP-glucuronosyltransferases (UGTs) and 15 ATP-binding cassette transporters (ABC transporters) were identified to be down-regulated in winged M. dirhodum individuals in the third instar nymphs, while 2 P450s, 2 CarEs, 1 UGT and 5 ABC transporters were up-regulated (Fig. 10A). In the fourth instar nymphs, 13 P450s, 10 CarEs, 6 GSTs, 13 UGTs and 6 ABC transporters were identified to be down-regulated in winged M. dirhodum individuals, while 3 P450s, 1 CarE, 1 UGT and 6 ABC transporters were up-regulated (Fig. 10B). In the adults of M. dirhodum, 10 P450s, 10 CarEs, 7 GSTs, 18 UGTs and 6 ABC transporters were identified to be down-regulated in winged individuals, while 1 P450, 2 CarEs, 1 UGT and 5 ABC transporters were up-regulated (Fig. 10C). Among them, 11 P450s, 12 CarEs, 7 GSTs, 15 UGTs and 15 ABC transporters were down-regulated in winged M. dirhodum individuals in all the three sampling stages (the third, fourth instar nymphs and adults), while 1 CarE, 1 UGT and 4 ABC transporters were up-regulated (Fig. 10).
More detoxification genes were highly expressed in wingless M. dirhodum. It seems that the wingless aphids may have stronger ability in detoxification metabolism. Considering that wingless aphids mainly harm host plants in their habitats, these highly expressed detoxifications might be important for them to resist the effects of plant secondary metabolites and insecticides. Actually, several of these genes have already been determined to be involved in insecticide resistance. For example, CYP6DC1 was significantly up-regulated in acetamiprid resistant strain of Aphis gossypii. RNA interference-mediated knockdown of CYP6DC1 significantly increased the sensitivity of resistant strain to acetamiprid54. mRNA level of GSTs2 was significantly higher in a field resistance population in Cydia pomonella, and further metabolic assays indicated that λ-cyhalothrin could be depleted by recombinant GSTs255. Although the number of highly expressed detoxification enzyme genes in winged M. dirhodum is much less than that of winged individuals, several of these genes have also been confirm to be important in insecticide resistance. For example, the sensitivity of Aphis craccivora to imidacloprid was significantly increased after knockdown of CYP380C6 by RNAi56. CYP380C6 and CYP6CY21 were overexpression in cyantraniliprole resistant strain of A. gossypii. Transgenic expression of CYP380C6 and CYP6CY21 in D. melanogaster suggested that they were sufficient to confer cyantraniliprole resistance, and also related to α-cypermethrin cross-resistance57.
Studies have pointed out that macropterous N. lugens was more susceptible to the neonicotinoid insecticides than that of the brachypterous ones. On the contrary, brachypterous N. lugens was more susceptible to the organophosphorus insecticide chlorpyrifos than the macropterous ones. While no significant difference in the susceptibility to nitenpyram, cycloxaprid, dichlorvos, buprofezin, isoprocarb, pymetrozine and etofenprox was observed between macropterous and brachypterous N. lugens58. We speculated that both winged and wingless insects need to resist insecticides and other toxics in the process of feeding and migration, and diverse detoxification enzymes might be adopted in winged and wingless M. dirhodum. The identification of these DEGs is very helpful to explore the different mechanisms of detoxification metabolism between winged and wingless aphids.
{kind=link}