Fabrication and optimization of LCHNPs. In this study, LCHNPs were prepared using a simple emulsification technique. LCHNPs could be thought to consist of a polymeric PLGA core surrounded by a layer of lipid, DOTAP. By varying the DOTAP-to-PLGA ratio, LCHNPs with various EE, LE and ζ-potentials could be achieved (Table 1).
From Table 1, several observations could be drawn on the effects of the masses of PLGA and DOTAP used on the characteristics of the resulting LCHNPs. A comparison between LCHNP60:10 and LCHNP120:10 highlighted that a higher quantity of PLGA used during fabrication was associated with an increase in vancomycin EE, and a decrease in LE and ζ-potential. The increase in EE from 26.12–44.19% with higher PLGA quantity was consistent with several published studies. These studies stated that an increase in PLGA quantity used during fabrication led to an increase in the core size of the nanoparticles, allowing for more vancomycin molecules to be encapsulated [14, 15]. A decrease in LE from 49.34 µg/mg to 40.79 µg/mg was observed when a higher quantity of PLGA was used. With an increase in the nanoparticle core size, there is also an increase in the mass of each NP, which likely explains the reduction of LE since LE is inversely proportional to the mass of nanoparticles. Also, the stronger negative ζ-potential of -26.37 mV for LCHNP120:10 as compared to -9.58 mV for LCHNP60:10 was due to the higher quantity of PLGA used in LCHNP120:10. This suggested that there was insufficient DOTAP to coat the PLGA core adequately. The EE and LE of LCHNP120:20 were reduced when compared to LCHNP60:20; this appeared to be an anomaly as the EE was expected to increase when more PLGA was used. LCHNP120:20 was visually observed to be a sticky paste instead of a white powder like the other samples; it was postulated that the high quantities of PLGA and DOTAP used during the fabrication of LCHNP120:20 led to an increased viscosity of the overall emulsion, and thus resulted in poor fabrication of the nanoparticles.
A comparison between LCHNP60:10 and LCHNP60:20 revealed that the higher mass of DOTAP used in LCHNP60:20 was associated with an increase in EE, LE and ζ-potential. Baek et al. suggested that hydrophilic drugs, such as vancomycin, tend to localize on the surface of hydrophobic PLGA nanoparticles. By incorporating a layer of lipid onto PLGA nanoparticles, hydrophilic drugs could be trapped between the PLGA core and lipid layer, resulting in increased EE and LE [7]. Thus, in the case of LCHNP60:20, a higher quantity of DOTAP used could lead to a more robust lipid layer, as verified by the strong, positive ζ-potential. Consequently, the robust layer of DOTAP contributed to the trapping of hydrophilic vancomycin molecules within the LCHNP, resulting in higher EE and LE.
Amongst LCHNP60:10, LCHNP 60:20, LCHNP120:10, and LCHNP120:20, it was observed that LCHNP60:20 displayed desirable properties required for the delivery of vancomycin into biofilms, whereby: 1) a high value of LE signifying that more antibiotics could be delivered with smaller quantity of LCHNPs; 2) a high value of EE indicating that a higher proportion of antibiotic used during fabrication was encapsulated, resulting in lesser wastage; and 3) a positive surface charge which would be beneficial for penetrating into biofilms. Thus, further optimization was carried out based on the formulation for LCHNP60:20, which led to LCHNP45:20 and LCHNP30:20.
It was noted that LCHNP45:20 had the highest EE, LE and ζ-potential of 48.66%, 72.99 µg/mg and +36.13 mV respectively (Figure 1a-c). These measurements were significantly higher than those of LCHNP60:20 and LCHNP30:20. In fact, the LE of the optimized LCHNP45:20 were significantly higher than the widely reported values of 2.6 – 15.8 µg/mg for reported nanoparticulated systems used to encapsulate hydrophilic molecules [16–20]. Thus, the formulation for LCHNP45:20 was chosen for subsequent experiments.
Physical characterization of optimized LCHNPs and comparison with PLGANPs. To determine the role of DOTAP in the nanoparticulate systems, bare and vancomycin-loaded PLGANPs were fabricated using the exact same parameters as those used for the fabrication of the optimized LCHNPs, less DOTAP. Through dynamic light scattering, the hydrodynamic diameters of both PLGANPs and LCHNPs were determined to be 221.03\(\pm\)6.50 nm and 207.83\(\pm\)2.06 nm respectively (Figure 2a). The sizes of these nanoparticles fall within the desirable range of 100 – 500 nm for penetration into biofilms through water channels and EPS without eliciting rapid clearance by the host [21]. LCHNPs were observed to be significantly smaller (**) than the bare PLGANPs. This was likely due to the presence of the cationic DOTAP, which played a crucial role in regulating the size of LCHNPs, preventing the coalescence of these nanoparticles. The drastic difference in ζ-potentials of PLGANPs and LCHNPs (-36.83\(\pm\)1.24 mV and 36.13\(\pm\)0.31 mv respectively) suggested that DOTAP was successfully coated onto the PLGA core of LCHNPs. The FESEM images (Figures 2b & 2c) showed the spherical morphologies of both PLGANPs and LCHNPs. The core-shell structure of LCHNPs was further verified using TEM (Figure 2d).
The EE and LE values of LCHNPs were significantly improved over those of PLGANPs. The addition of the DOTAP layer on the PLGA surface possibly prevented vancomycin molecules from escaping out of the nanoparticles during fabrication, leading to higher EE and LE. Given that vancomycin is highly hydrophilic, there is a high tendency of it escaping from the oil phase of the emulsion into the bulk aqueous phase during the fabrication process. This could be observed from the lower EE and LE values of PLGANPs of 24.11\(\pm\)0.64% and 52.25\(\pm\)1.38 µg/mg respectively. On the other hand, LCHNPs had an EE of 48.66\(\pm\)0.42% and LE of 72.99\(\pm\).63 µg/mg, highlighting the essential role of the lipid layer in achieving high EE and LE of hydrophilic molecules in polymeric nanoparticles [22, 23]. Furthermore, the lipid layer was observed to retard the release of vancomycin from the nanoparticles. From the release profiles (Figure 2e), 43.93% of the encapsulated vancomycin within Van-LCHNPs was released, while 92.26% of the drug was released from Van-PLGANPs. The burst release of vancomycin from both types of nanoparticles within the first 24 hours was not unexpected, given that the hydrophilic vancomycin would tend to localize at the surface of the hydrophobic nanoparticles. Nonetheless, vancomycin release data within the first 24 hours suggested that the presence of the lipid layer mitigated the burst release of vancomycin. This was in good agreement with Sivadasan et al. who suggested that the presence of a lipid shell on nanoparticulate carriers reduced the rate of water penetration, resulting in reduced rate of hydrolysis of the polymeric core and thus, a slower rate of release [23].
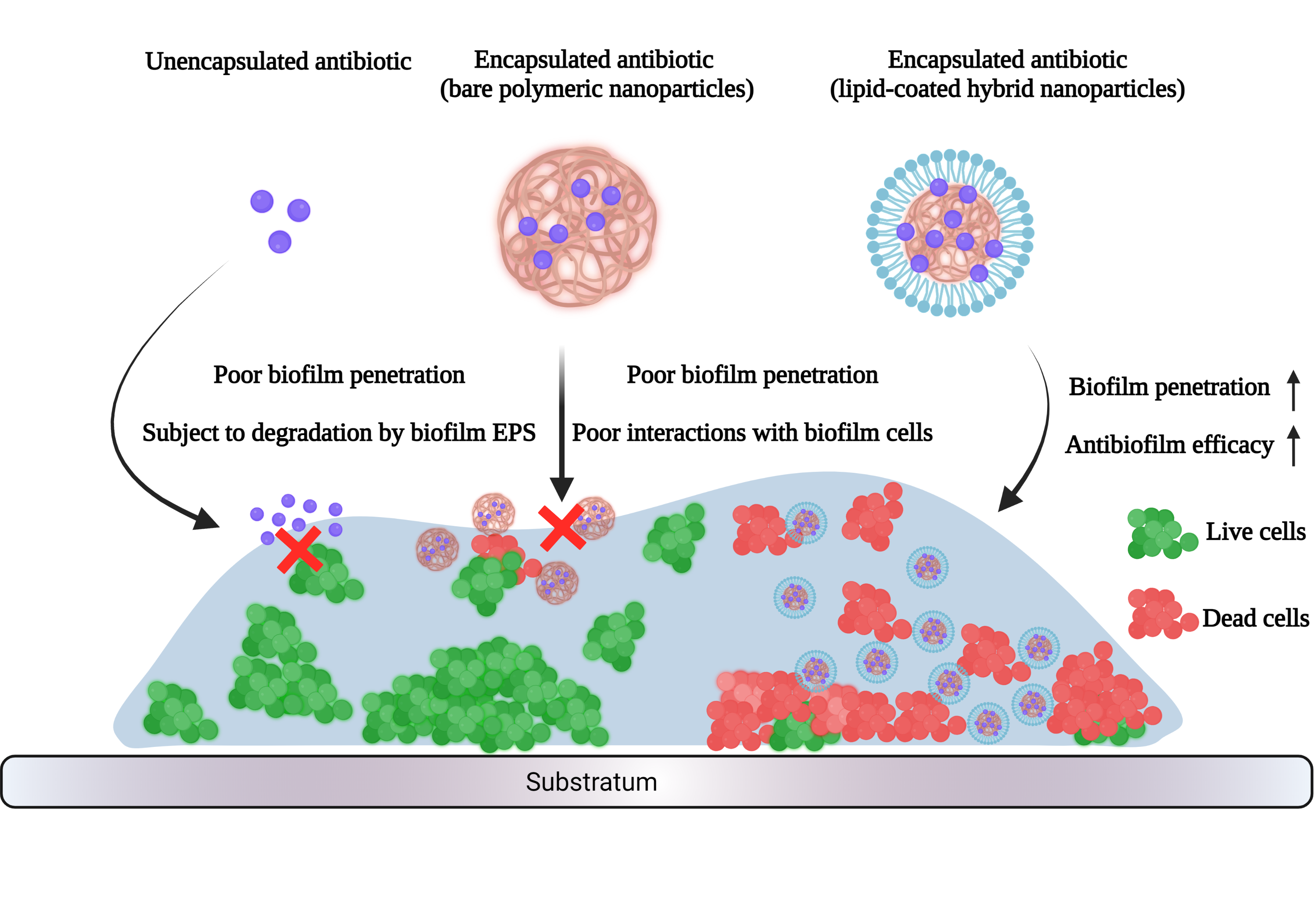
Presence of DOTAP layer enhanced antibacterial efficacy against planktonic USA300. Prior studies had demonstrated the effectiveness of positively-charged drug delivery systems, such as cationic liposomes and certain antimicrobial peptides, on making first contact and continuous binding with bacteria [24, 25]. Therefore, it was hypothesized that Van-LCHNPs, being positively charged, would have an enhanced therapeutic effect against planktonic USA300 compared to bare Van-PLGANPs. Comparing against Free-Van, it was observed that Van-LCHNPs exhibited enhanced antibacterial effects against planktonic USA300 (Table 2). By loading vancomycin into LCHNPs, there was a reduction in MIC and MBC by at least six-fold as compared to the MIC and MBC values for Free-Van. This contrasted with the case for Van-PLGANPs whereby MIC and MBC values increased. PLGANPs have poor interactions with the USA300 cells due to the negative charge on both. Arakha et al. reported that negatively-charged nanoparticles do not adhere to bacteria owing to poor electrostatic attraction [26]. Furthermore, the controlled release of vancomycin from Van-PLGANPs implied that inhibitory or biocidal concentrations of vancomycin could not be reached quickly, ultimately resulting in higher MIC and MBC values observed as compared to Free-Van. To eliminate the possibility that the enhanced bacterial effects of LCHNPs were caused directly by the intrinsic antibacterial property of the nanoparticles, blank LCHNPs and PLGANPs were utilized as controls; data from these blank runs showed that no antibacterial effects were observed from extremely high nanoparticle concentrations up to 10,000 µg/mL. The stronger antibacterial effect of Van-LCHNPs in comparison to Van-PLGANPs could be due to the presence of the cationic DOTAP layer playing a significant role in allowing the LCHNPs to make first contact and continuous binding with the USA300 cells. This is in good agreement with Costa et al. who correlated the positive effect of enhancing interactions between nanoparticles and bacterial cells on antibiotic delivery [27].
Enhanced antibiofilm effect against USA300 biofilms using LCHNPs. To evaluate the capability of LCHNPs to eradicate USA300 biofilms, static biofilms were cultivated in 8-wells chambered coverglasses. The cultivated biofilms were then treated overnight with Free-Van, Van-PLGANPs and Van-LCHNPs with vancomycin concentrations ranging from 256 µg/mL to 0.25 µg/mL in 4-fold dilutions. Through crystal violet staining, remnant biofilms following treatment could be visually observed (Figure 3a-c). It was observed that Free-Van and Van-PLGANPs did not cause any significant reduction in biomass across all tested concentrations as compared to the untreated controls. This suggested that both groups were not able to sufficiently eradicate the underlying bacterial cells to cause any biofilm disruption. On the other hand, significant reductions in biomass were observed for Van-LCHNPs, specifically for vancomycin concentrations of 256 – 4 µg/mL. It should be noted that vancomycin concentrations of 256 µg/mL and 64 µg/mL for Van-LCHNPs corresponded to higher concentrations of nanoparticles that were used for treatment. The high concentrations led to residual nanoparticles that were not completely removed during the washing step in the protocol which subsequently led to the observed signals following crystal violet staining which should not be incorrectly interpreted as biomass. The observations from crystal violet staining were supported with CFU data that were derived from remnant biofilms. As observed from Figure 3d, Van-LCHNPs exhibited an exceptional capability to eradicate USA300 biofilms across all tested concentrations of vancomycin. At 256 µg/mL, Van-LCHNPs showed the highest degree of biofilm cells eradication with 4 logs reduction (or 99.99%) in CFU/mL as compared to the untreated control (~ 2 x 1010 CFU/mL). Even at 0.25 µg/mL of vancomycin, Van-LCHNPs could reduce the CFU count by 2 logs (99%). On the other hand, Free-Van and Van-PLGANPs were largely incapable of meaningfully reducing the CFUs counts in USA300 biofilms. This suggested that Free-Van and Van-PLGANPs could not adequately penetrate through the biofilms for vancomycin to be uptake by the underlying bacterial cells. We postulated that the strong antibiofilm capability of Van-LCHNPs is due to the presence of the DOTAP coating which enhanced interactions between the nanoparticles and the biofilm EPS, leading to enhanced penetration. Wang et al. suggested that the utilization of lipids which are chemically similar to bacterial membranes could improve the fusion of nanoparticles with bacterial cells [28]. Furthermore, Sugano et al. reported that cationic liposomes do not only interact well with planktonic bacterial cells, but also biofilms [25]. This was due to electrostatic interactions between the cationic liposomes and biofilm matrix, which is often negatively charged. Consequently, Van-LCHNPs, coated with cationic DOTAP, were able to penetrate deeper into the biofilms to deliver the antibiotics directly to the USA300 cells.
Enhanced antibiofilm capability of LCHNPs could prevent recalcitrant infections. A key challenge in treating biofilm infections arises from the fact that it is hard for antibiotics or antibiotic delivery system to penetrate deeply into biofilms to eradicate the underlying cells uniformly throughout the entire thickness of the biofilm [28]. Even if killing occurs, biofilm cells may not be sufficiently eradicated, and the biofilm matrix remains intact. This often gives rise to recalcitrant infections as biofilms which are not completely removed often become a starting point for biofilm regrowth [29]. From Figure 3c, it was observed that there were little biomass remaining following treatment with Van-LCHNPs corresponding to 4 – 256 µg/mL, suggesting that biofilms were removed. To verify this, confocal microscopy was performed following treatment to image the remaining biofilms. The biofilms were stained with Syto9 and PI prior to imaging.
Figure 4a shows the confocal microscopy images for each sample group. When biofilms were treated with Free-Van (0.25 – 16 µg/mL), the confocal images showed that the bulk of the biofilm cells were alive (green signals) even though the tested concentrations exceeded the MBC of vancomycin required to completely eradicate planktonic USA300 cells. Also, digital measurement of the biofilm thicknesses (Figure 4b) showed that biofilms treated with Free-Van had increased in thickness as compared to the untreated control. The increase in biofilm thickness could be due to the mechanism of action for vancomycin, by interrupting cell wall synthesis resulting in bacterial lysis. As a result, biomolecules leaking from the lysed cells may be retained in the biofilm, leading to an increase in biofilm thickness. For Van-PLGANPs, only at concentrations corresponding to 4 µg/mL and 16 µg/mL of vancomycin exhibited any antibacterial effect towards the biofilm cells. The killing of biofilm cells at 4 µg/mL appeared to be closer to the top surface of the biofilm, while biofilm cells residing deeper remained alive. At 16 µg/mL, the biofilm cells appeared to be eradicated throughout the thickness of the biofilm. However, the biofilm remained intact and could lead to regrowth of the biofilm. For Van-LCHNPs, the thickness of the biofilms appeared to be significantly reduced across all 4 concentrations that were imaged. The degree of biofilm removal was most pronounced at concentrations of 1 µg/mL, 4 µg/mL and 16 µg/mL. Antibiofilm effects appeared to be dose-dependent, whereby the dead cells appeared to be distributed throughout the remaining thickness of the biofilms for 4 µg/mL and 16 µg/mL. Overall, the data suggested that Van-LCHNPs possessed excellent antibiofilm ability as it could eradicate the underlying cells and cause biofilm removal.
Presence of a robust coating of DOTAP led to enhanced penetration ability of LCHNPs into USA300 biofilms. It was verified through several methods that Van-LCHNPs possessed exceptional antibiofilm ability over Free-Van and Van-PLGANPs (Figures 3 and 4). This study stemmed from the hypothesis that the presence of a cationic lipid coating could enhance the penetration of nanoparticles into biofilms, leading to improved antibiofilm capabilities. To verify role of DOTAP coating in enabling enhanced penetration into biofilms, PI-loaded LCHNPs of various DOTAP:PLGA ratios (Figure 5a) were fabricated. These PI-loaded nanoparticles were then incubated overnight with USA300 biofilms prior to confocal microscopy imaging. By utilizing the Z-stack function of the confocal microscope, it was investigated how deep into the biofilms these nanoparticles were able to penetrate.
Confocal images (Figure 5) showed that nanoparticles with DOTAP:PLGA ratio of 0:45 (equivalent to PLGANPs) coating exhibited poor penetration into USA300 biofilms while penetration into biofilms increased gradually as the DOTAP:PLGA ratio was increased from 0:45 to 20:45. This could be observed from the increase in red signals coming from the PI-loaded nanoparticles as the DOTAP:PLGA ratio was increased. This suggested that DOTAP:PLGA ratio and the degree of penetration into biofilms had a dose-response relationship, implying that the DOTAP coating played a causal role in allowing the LCHNPs to penetrate deeply into the biofilms. To the best knowledge of the authors at the time of writing, this is the first time that a cationic lipid coating is shown to directly enhance the penetrating capability of nanoparticles, especially for LCHNPs, into biofilms.
The penetration of LCHNPs into biofilms could be described using the nanoparticle-biofilm transport phenomena. Firstly, nanoparticles must be transported to the biofilm-fluid interface; secondly, the nanoparticles must attach to the outer surface of the biofilms; lastly, the nanoparticles would have to migrate into the biofilm. Interactions with the biofilm EPS are often governed by the surface properties of the nanoparticles, such as surface charge and functional groups present on the surface. These interactions are critical for nanoparticles to establish first contact (attachment) required with the biofilm surface before the nanoparticles could migrate within the biofilms [30]. In the case of LCHNPs in this study, a strong positively charged surface was presented to the biofilm EPS, allowing for strong interactions between the negatively charged EPS components and the LCHNPs. This contrasts with PLGANPs, which present a negatively charged surface and thus, have poor interactions with the biofilm EPS. This observation is congruent with studies that reported that electrostatic interactions played a critical role in determining the ability of nanoparticles to adhere onto biofilm surfaces [31, 32].


{kind=link}