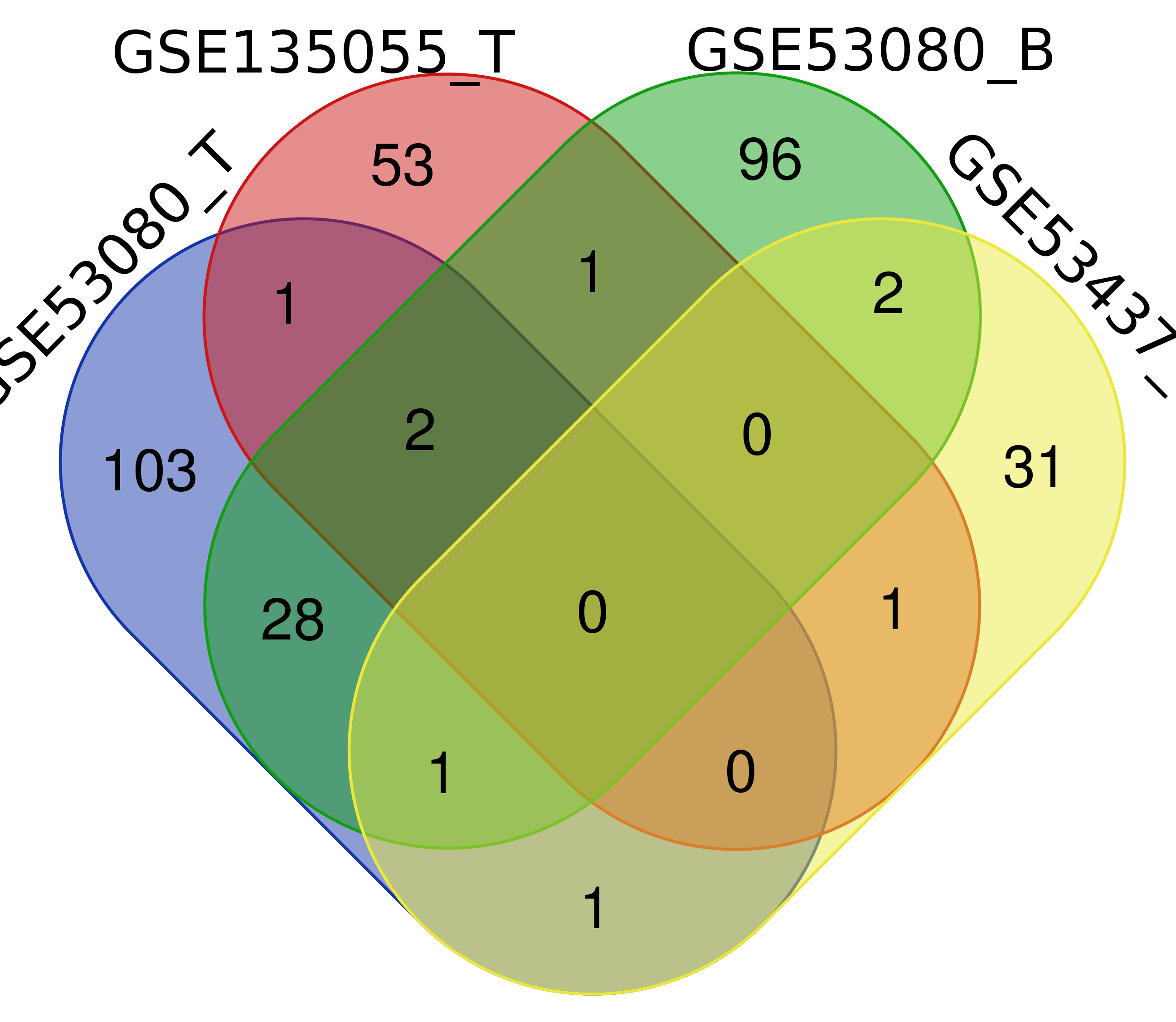
This study identified 5upregulatedDEmiRNAs (hsa-miR-216a-3p, hsa-miR-216a-5p, hsa-miR-217-3p, hsa-miR-217-5p and hsa-miR-208a) in myocardial tissue and blood fromthemiRNA profiles of GSE135055, GSE53080, and GSE53437 and extracted consistentmiRNAswith multifold higher differential expression to distinguish HFrEF patients fromthecontrol population.miR-217 was also verifiedupregulatedinamouse model of heart failure caused by ischemia and hypertrophy.
Hsa-miR-216a and hsa-miR-217 screened in this study have also been verified by previous studies screening circulating miRNAs as potential biomarkers for HF9. This study showed thatthecombination of miR-216a and miR-217 or more miRNAs could be considered a new diagnostic biomarker for HF.Previous studies have also shown increased expression of miR-217 in the hearts of CHF patients and mice with TAC surgery10. Our findings are consistent with these findings. Overexpression of miR-217 in vivo aggravated cardiac hypertrophy, fibrosis, and cardiac dysfunction caused by stress overload10, 11. In addition, miR-217 wasupregulatedin ischemia-reperfusion model mice and in hypoxia/reoxygen-treated H9C2 cells12.Dual-specificity protein phosphatase 14 (DUSP14) has been shownto bea target, inactivatingtheNF-κB and MAPK pathways to prevent MI reperfusion injury12. Therefore, miR-217 could be a promising therapeutic target in HF cardiac hypertrophy mechanisms.
To date, miR-208has beenreportedto be amyocardium-specific miRNA7, 13. Significant upregulation of miR-208a and related proteins was also presented in human hearts with type 2 diabetes14. However, its high expression was not observed in the myocardium of patients with HFrEF in the 3 profiles we included, but it was highly expressed in the blood samples, which was also verified intheplasma of HF patients in another study15.Together, no current evidence has shown miR-208a asaDEmiRNA in the myocardial tissue of MI and TAC mouse models,and this study also confirmedthis observation. However, miR-208a silencing can attenuate myocardial apoptosis and promote cardiac repair16, which needs further exploration.
Regarding the manifestation of the potential biological roles of 5 miRNAs in human heart failure, 9 key genes were enriched and analyzed, most of which were confirmed to be related to HF. JAK-STAT signaling is associated with cardiac hypertrophy and remodeling induced by pressure overload and ischemia-reperfusion-induced cardiac dysfunction17. SOS1, PTPN11, and KRAS mutations may be associated with hypertrophic cardiomyopathy of RASopathy18. PIK3R1 is modulated bytheFoxO3a pathway,which is highly phosphorylated in dysfunctional hearts19. However, there are no relevant reports on the role of UBE2N (E2 ubiquitin-conjugating enzymes) in HFrEF and even in the heart. This study was the first toidentifyUBE2N not only as a hub gene but alsoasa key molecule in MCODE module 1, which needs to be explored further.
Gene enrichment analysis revealed that several signaling pathwayswereinvolved in the regulation oftheTGF-β signaling pathway, the FoxO signaling pathway, theToll-like receptor signaling pathway, stress response, and hormone signaling pathways,such as those for thyroid and steroid hormones, which arewidelyreported to correlate with HF20-23. Additionally, the signaling molecules in each pathway may becorrelated.FoxO3a has been shown to act as a negative regulator of TGF-β1-induced cardiac extracellular matrix (ECM) transformation24, and TLR2 deficiency leads to excessive activation of the transcription factor Forkhead box protein O1 (FoxO1), which affects age-related cardiac function in mice25.TLR4 activates the expression of severalproinflammatorycytokine genes in myocardial inflammation26.
Our study has some limitations. First, the etiology of heart failure is not completely presented in this study,andonly the common etiology is constructed.Second, we did not choose to use mouse serum for verification.This isbecauseprevious studies have shown thatmiRNAsobserved in human patients with HF are mostly present at low levels and cannot be validated in well-established mouse models27.
This study may shed new light on the combination ofbiomarkersin HF by combining multiple expression profiles.Multi-miRNAs combined with or without BNP still significantlyimprovethe prediction model to distinguish heart failure with preserved ejection fraction (HFpEF) from HFrEF andimprovespecificity and accuracy28.Therefore, with increased sample size and reduced deviation of detection, this study ultimately improved the authenticity of the findings. In addition to preliminary verification at the animal level, the miRNA screened is expected to be used asapanel in the next step to distinguish HFrEFs in our center.
In conclusion, we identified valuable biomarkers in tissues and blood and established a miRNA-mRNA regulatory network. In addition, bioinformatics techniques were applied to explore the crucial molecules involved in the development of HFrEF. We found that miR-217 was consistently expressed in HFrEF through animal heart failure models, offering attractive potential as a biomarker and therapeutic target for heart failure.
{kind=link}