Genome-Wide Identification of Chitinase Genes and Their Chromosomal Distribution.
We explored VvChis in grape from the whole genome sequence scaffolds of ‘Piont Nior’. HMMER 3.0 and the Pfam protein family databases with the GH18 and GH19 domains were used to search for chitinases in grape genomes. SMAT and INTERPROSCAN programs were used to vertify the predicted genes. As a result, we identified 42 VvChis in grape (Supplementary Table S2). Among them, VvChi1 ~ VvChi25 were searched out based on GH18 domains, and VvChi26 ~ VvChi42 were searched out based on GH19 domains.
The 42 identified VvChis were found in 9 out of the 19 grapevine chromosomes, as well as in the unplaced contigs chromosome (ChrUn), which contains sequences whose physical position on specific chromosomes have not yet been defined. As shown in Fig. 1, fourteen, seven, seven, four, three, two, one and one chitinase genes located at the chromosomes 5, 11,15, 6, 4, 16, 14 and 3, respectively. Four chitinase genes were also found in ChrUn.
Figures, Tables and Schemes
The phylogenetic relationships of VvChis were further analyzed by constructing an unrooted tree using the maximum likehood method. The results indicated that the 42 VvChis were clustered into five groups, namely A, B, C, D and E (Fig. 2a). These classifications were consistent with the presence of conserved and catalytic domains (Fig. 2b), and exon-intron organizations (Fig. 2c). Twenty-five chitinases belonging to GH 18 family were mainly divided into groups A, B and E, except VvChi24. Seventeen members of the GH 19 family were classified into groups C and D.
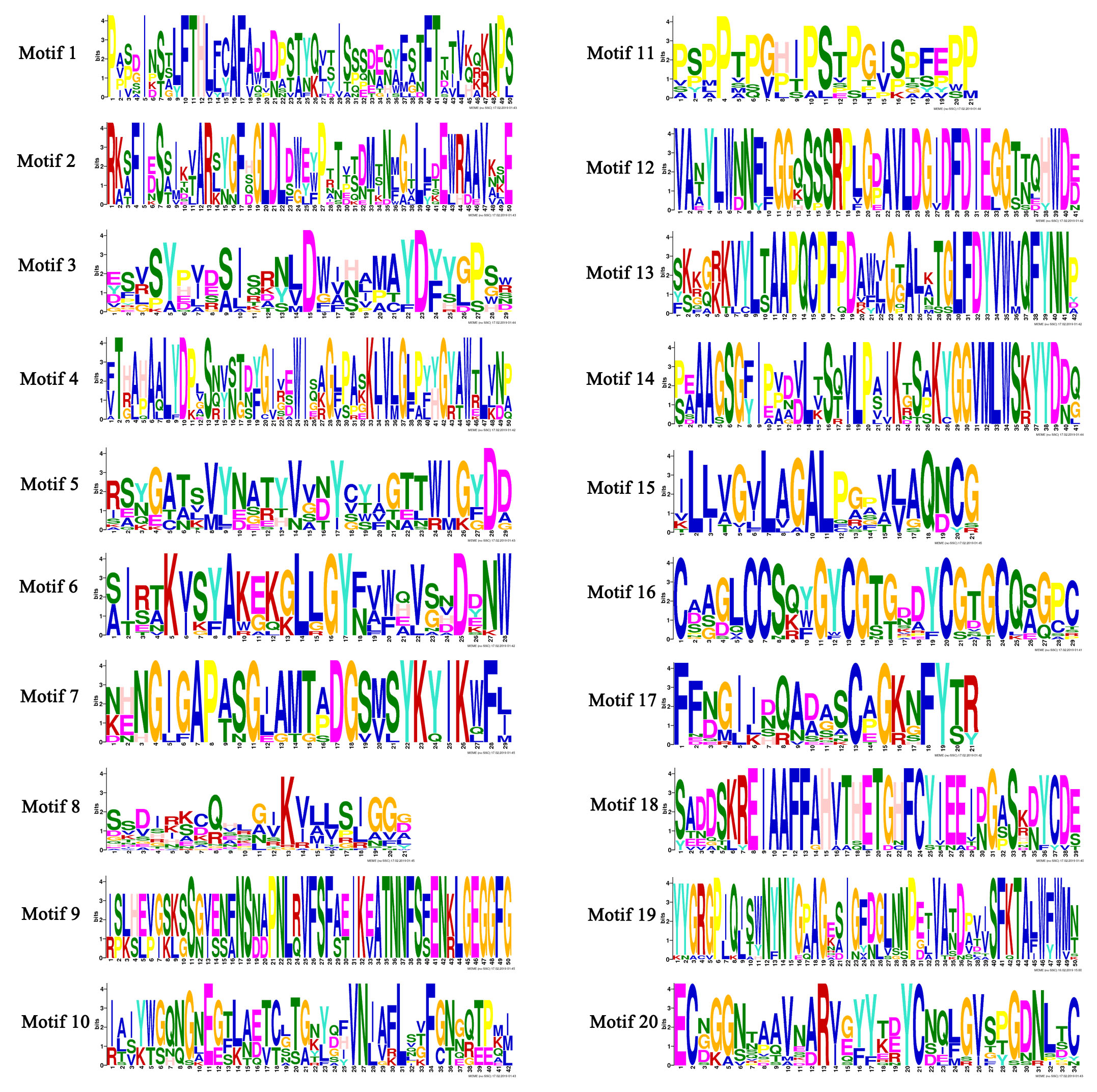
The conserved motifs of chitinases were displayed schematically in Fig. 2b. Twenty conserved motifs in the 42 VvChis were analyzed (Figure S1). The motifs 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 and 14 were found in the VvChi belonging to GH18 genes family while the motifs 15, 16, 17, 18, 19 and 20 were only detected in those of GH19 family. The type, number, order and motif location of these chitinase were similar within each group, but significantly different from others. The members in group A displayed the same five motif components (motif 8, 10, 12, 13 and 14) ranking in an identical order. The same situation was also found in group C (15, 16, 17, 18, 19 and 20) and 80% in group E (1, 2, 3, 4, 5, 6 and 7). The members in group B homogeneously shared motif 8. VvChis in Group C and D contained the same motifs (15, 16, 17, 18, 19 and 20) but with different number which indicated that these two group had a closer evolutionary and phylogenetic relationship. These findings indicated that the motif compositions of each group in chitinase genes family were relatively conserved. Through the analysis of protein secondary structure (Table S3), all VvChis were with four structures, namely Alpha helix, Extended strand, Beta brige, and Random coil, among which the proportion of Alpha helix and Random coil was the highest, while that of Beta brige was the lowest. The protein secondary structure had no significant disparity in different groups.
From the gene structure of VvChis (Fig. 2c), VvChi34 was longest, and VvChi9 was the shortest. Most genes in the same group generally shared exon-intron organizations with the same number of exons or introns. For example, one or two exons were found in most genes of group A, B and E while at least two exons existed in genes of group C and three in group D. These results showed that the VvChis in GH 19 had more exons than that in GH 18. Overall, the results of the conserved motifs location/components, and exon-intron gene structure of the chitinases in grape were consistent with the phylogenetic classification as well. Moreover, the highly conserved sequences of chitinase family members in the same group indicated that these genes were subject to duplications during their evolution.
Cis-Elements Analysis in the Chitinase Genes Promoters and Functional Prediction of Chitinases
To further clarify the gene function and transcriptional regulation mechanism of VvChis, we amplified the 1500 bp sequence upstream of VvChis and analyzed their cis-elements. These predicted cis-elements related to various process including photoreaction, hormone responsiveness, biotic and abiotic stress responsiveness, and plant development differed among groups and genes (Fig. 3). In the promoter of all genes, the number of elements related to endosperm expression was the largest except those related to light response, indicating that chitinases might had relationship with seeds development.
Compared with other groups, group A had the largest number of cis-element types, but B had the least. In the promoter of group B, there were many elements for gibberellin-responsiveness (GARE), but no for ethylene-responsiveness (ERE). Specifically, would-responsive elements were only found in the promoter of group C and fungal elicitor related elements located in the promoter of all groups of VvChis. Among 42 VvChis, VvChi27 had the largest number of cis-elements. Dehydration, low-temp, salt stresses related elements were only detected in the promoter of VvChi10, while cold- and dehydration-responsiveness related elements were only found in VvChi39, and cell cycle regulation elements only in VvChi10 and VvChi17. Additionally, different elements constitute was also found even in the promoter of the same gene. For example, different from other members in group B, meristem expression and low-temperature responsiveness related elements were only found in the promoter of VvChi21 while abscisic acid responsiveness and drought-inducibility related elements only in VvChi20 and MeJA-responsiveness related elements only in VvChi23.
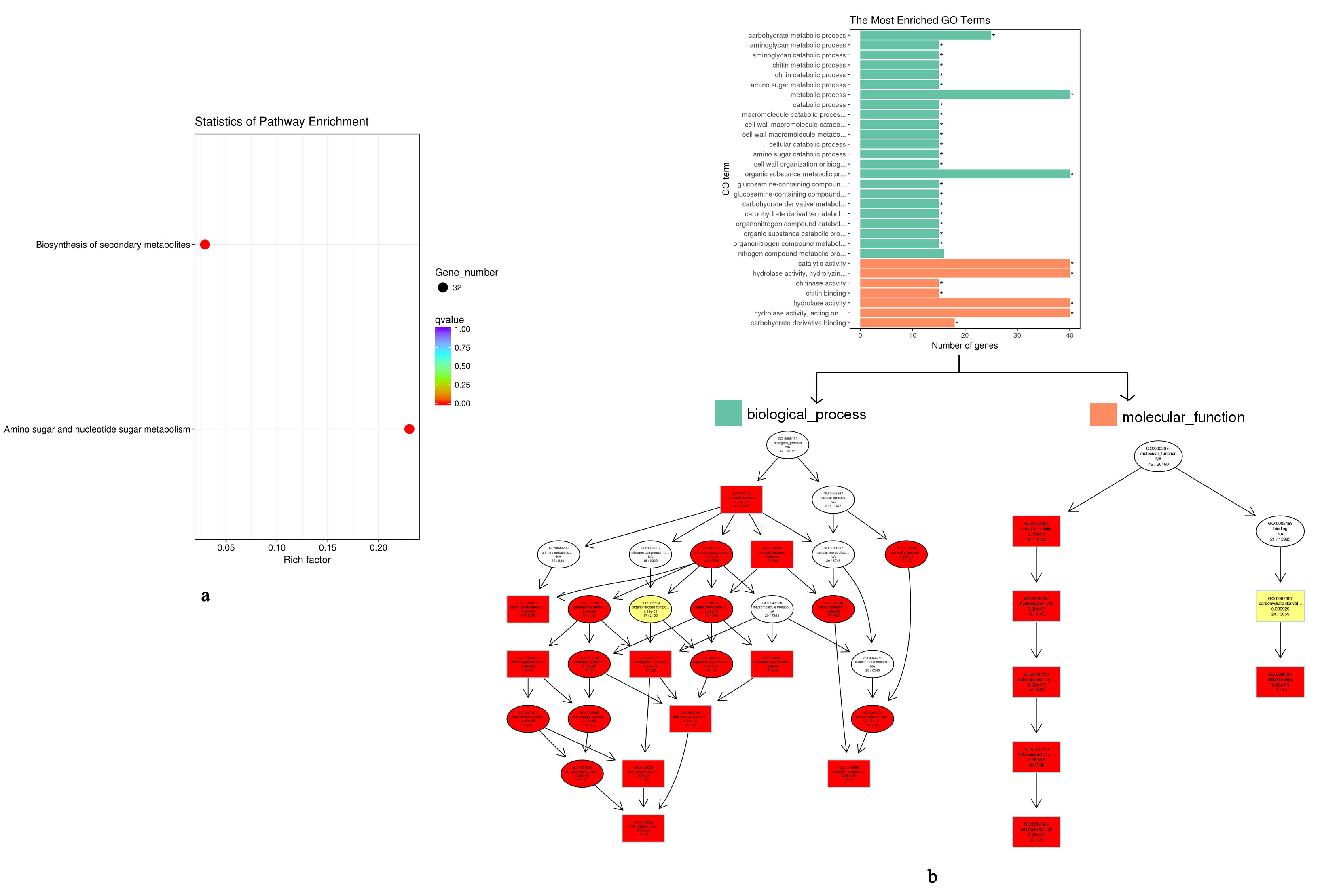
GO and KEGG analysis indicated that VvChis mainly participate in the biosynthesis of secondary metabolites and amino sugar and nucleotide sugar metabolism (Figure S2a). The biological process of VvChis were all attributed to cell wall macromolecule catabolic process and chitin catabolic process, while molecular function were attributed to chitin binding and chitinase activity(Figure S2b).
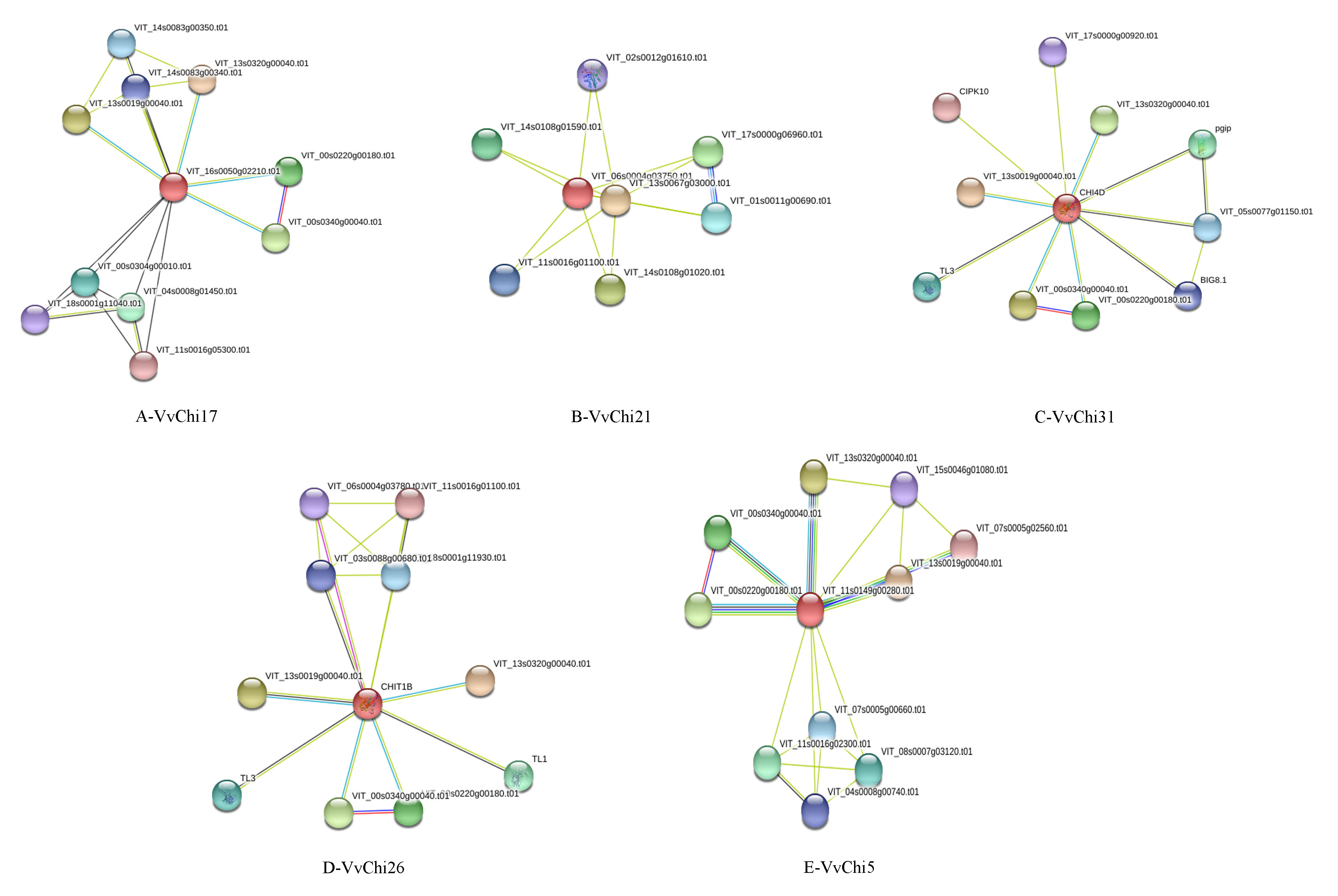
As we all know, chitinases play a major role in defense against fungal pathogens by directly attacking divading fungal pathogens. However, the specific regulatory network and effects on grapes is still not clear. Chi31 from group C could interact with PGIP and BG to participate in plant resistance to phytopathogenic fungi (Figure S3). Chi5 from group E, Chi17 from group A, Chi26 from group D, Chi21 from group B, and Chi31 from group C also interacted with sugar metabolism proteins to regulate plant growth, such as HEXO, UGDH, GAD, etc. In addition to the known sugar metabolism and disease-resistant related proteins, Chis could also interact with many undefined functional proteins, and their metabolic network remain to be further explored (Table S4).
Subcellular Localizations of Chitinase Genes
VvChi17, VvChi22, VvChi31, VvChi26 and VvChi5 were cloned and made a fusion protein with GFP, individually. The five VvCHI-GFP fusion proteins (VvChi7-GFP, VvChi22-GFP, VvChi31-GFP, VvChi26-GFP, and VvChi5-GFP) and the GFP control were transformed tobacco leaf epidermal cells. The fluorescent signal in tobacco leaves was detected with confocal laser scanning microscopy. As shown in Fig. 4, VvChi17 and VvChi5 located on plastids, VvChi22 and VvChi26 located on plasma membranes, and VvChi31 located on nuclear and plasma membranes.
Expression Analysis of Chitinase Genes in Grape
The expression level of twenty-five genes selected from five groups in different grape tissues was detected. There were differences in the expression profiles of VvChis genes in the seed cultivar ‘Kyoho’ and seedless cultivar ‘Summer Black’ (Fig. 5). Overall, the expression level of VvChis in various organs of ‘Kyoho’ was much higher than that of ‘Summer Black’, except tendril. The expression level of most genes in root was the highest and much lower in stem and flowers in ‘Summer Black’. Similarly, the highest expression level of most genes was found in ‘Kyoho’ root and the lowest expression level was found in tendril. It was worth noting that the expression level of most genes in seeds of ‘Kyoho’ was relatively higher compared to other tissues.
The expression level of VvChis from different groups was also different. In these two grapes’ root, the expression level of genes in group A, B, and E was higher than that in group C and D. In the skin, the transcription level of genes in group C and D was high while those in group A, B, and E hardly expressed. Similar to the genes in the skin, the expression of genes in group C and D was also in a high level. In the flowers, only the expression of VvChi31 and VvChi37 were expressed in a high level. In the seeds of ‘Kyoho’, the expression level of genes in group D was significantly higher than those in other groups.
Although the structure of VvChis was similar in the same group, the expression level of these genes was still different. In ‘Summer Black’, the expression level of VvChi22 in group B was higher than that of other members. VvChi16 in group A expressed lowly in root, stem, leaves, flower, flesh and skin, but showed higher expression level in tendril than any other genes. In group C, VvChi33 expressed highly in root, stem, leaves and tendril, while VvChi31 showed the highest expression level in flesh and skin, and VvChi37 highly expressed in flower. In ‘Kyoho’, VvChi12 expressed much higher than other members in group A. In group B, the expression level of VvChi25 and VvChi22 was significantly higher than that of other members. The expression level of VvChi31 was very low in the root, in which VvChi29, VvChi33 and VvChi34 in group C showed a high expression level; Additionally, the expression level of VvChi26 and VvChi27 in leaves was much higher than other genes in group D. VvChi5 and VvChi8 showed the highest expression level in stem, leaves, and flower, while other genes in group E showed the lowest expression level.
To investigate the spatial and temporal expression patterns of VvChis genes in grape, a global transcriptomic atlas comprising 54 tissues in different developmental stages was analyzed. Almost all VvChis genes had the corresponding probes on the NimbleGen array except VvChi8, VvChi25, VvChi27 and VvChi28. As a whole, the expression of VvChis showed the tissue–specific pattern (Fig. 6). Overall, the genes in group A, B and D showed a relatively low expression level, while those in group C and E remained at higher expression levels. The genes in different groups present the different expression patterns. For example, the genes in group C showed higher transcription level in berry pericarp, berry flesh, berry skins, flower, leaf, root and rachis, while those in group E mainly presented higher expression levels in bud, but lower in berry flesh, rachis and berry skins.
Effect of Botrytis cinerea Treatment on the Expression Pattern of VvChis
The expression level of VvChis was examined after 5 days of inoculation of grape leaves and berries with Botrytis cinerea (Fig. 7a). The results showed that most groups of VvChis were able to respond to pathogen infection (Fig. 7b).
Although the expression level of VvChis in leaves was significantly higher than that in berries, the response of berries to pathogens was much more sensitive than that of leaves. The expression level of these genes in the berries was 3 to 80 times higher than that in leaves. After the pathogen infects the leaves and berries of the grapes, the expression levels of the genes in the A, B and E groups showed a significant upward trend. From the degree of up-regulation, VvChi5 in group E, VvChi25 in group B, and VvChi11 (leaf) and VvChi16 (fruit) in group A were the most up-regulated. VvChi17 in group A, VvChi22 in group B and VvChi5 in group E had the highest expression levels in berries. The expression patterns of genes in group C and group D were different from those in other groups. The expression levels of VvChi37, VvChi29 and VvChi33 in group C increased after infection while VvChi31 and VvChi34 showed a downward trend. The expression levels of VvChi24, VvChi26, VvChi27 and VvChi28 in group D showed a downward trend after infection while the expression of VvChi42 showed an upward trend. Among these berries, the expression level of the VvChi31 in group C and VvChi24 in group D were the highest.
Interaction Vertification between the Chitinase and Metallothionein (MTL) or Mannose-binding lectin (MBL)
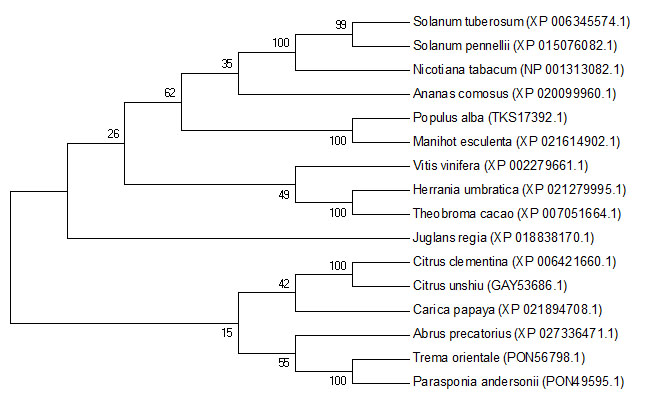
According to the phylogenetic analysis, VvChi17 from Vitis vinifera and Herrania umbratica, Theobroma cacao were clustered together with a far kindship to the others (Figure S4).
The vectors pGBKT7-Chi17, pGADT7-MTL and pGADT7-MBL were constructed. Then, the constructed expression vector pGBKT7-Chi17 and pGADT7-MTL or pGADT7-MBL were simultaneously transformed into yeast competent cells Y2H-Gold for interaction verification, individually. The results showed that the pGBKT7-Chi17 and pGADT7-MTL co-transfected yeast cells and the positive control could grow normally and turn blue, while pGBKT7-Chi17 and pGADT7-empty co-transfected and pGADT7-MTL and pGBKT7-empty co-transfected yeast cells had no interaction signal, indicating that the chitinase could interact with MTL. The yeast cells co-transfected with pGBKT7-Chi17 and pGADT7-MBL were able to grow normally but did not turn blue, which proved that pGBKT7-Chi had no interaction with pGADT7-MBL (Fig. 8).
To further verify the interaction between MTL and Chi17, we performed BiFC test. The yellow fluorescence was detected in tobacco epidermal cells only when YNE-MTL and YCE-Chi co-expressed (Fig. 9), further verifying the interaction between Chi17 and MTL.
Effect of VvChis on Disease Resistance of Tomato and Strawberry
To investigate the function of VvChis, we analysis the effects of VvChis overexpression on fruit ripening and disease resistance by transient expression during strawberry and tomato color-changing stage.
The results showed that the overexpression of VvChis could reduce the probability of tomato infection (Fig. 10), among these VvChi17 and VvChi31 were reduced by 42.9% and 64.3% respectively, with the disease index below 30.Considering with various physiological indicators (Table S5, S6), the ability on disease resistance of VvChis was: VvChi31 > VvChi17 > VvChi22 > VvChi5 > VvChi26. Different from tomato, although 5 VvChis can also promote disease resistance in strawberry, VvChi31 has the weakest disease resistance with the disease index 92.8%, and VvChi17 has the strongest ability with infection rate reduced by 78.6%. In conclusion, the function of VvChis in tomato and strawberry was significantly different, and VvChi17 exhibits strong resistance to disease in both two species.
Effects of hormones and environment stress on VvChis
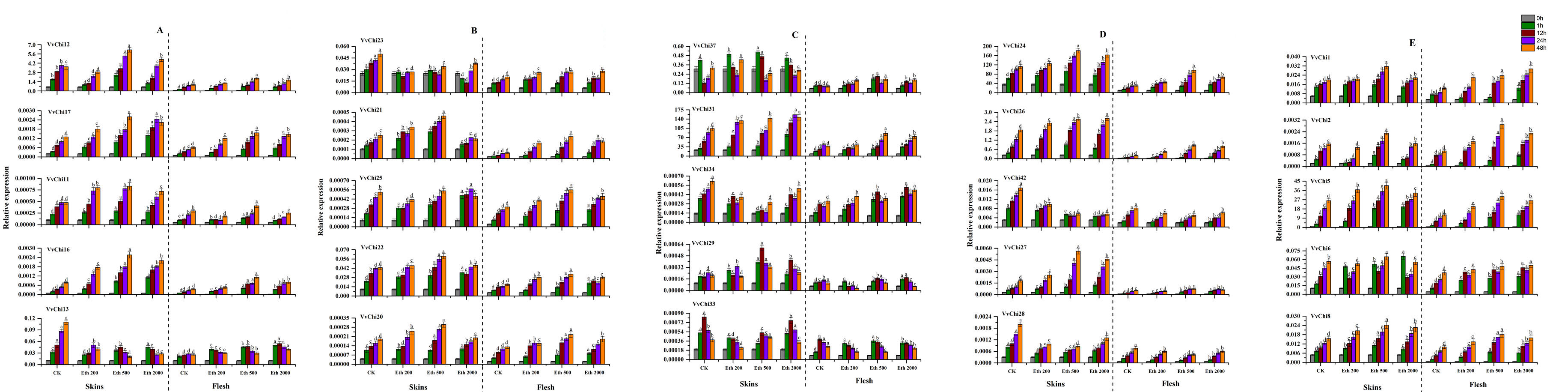
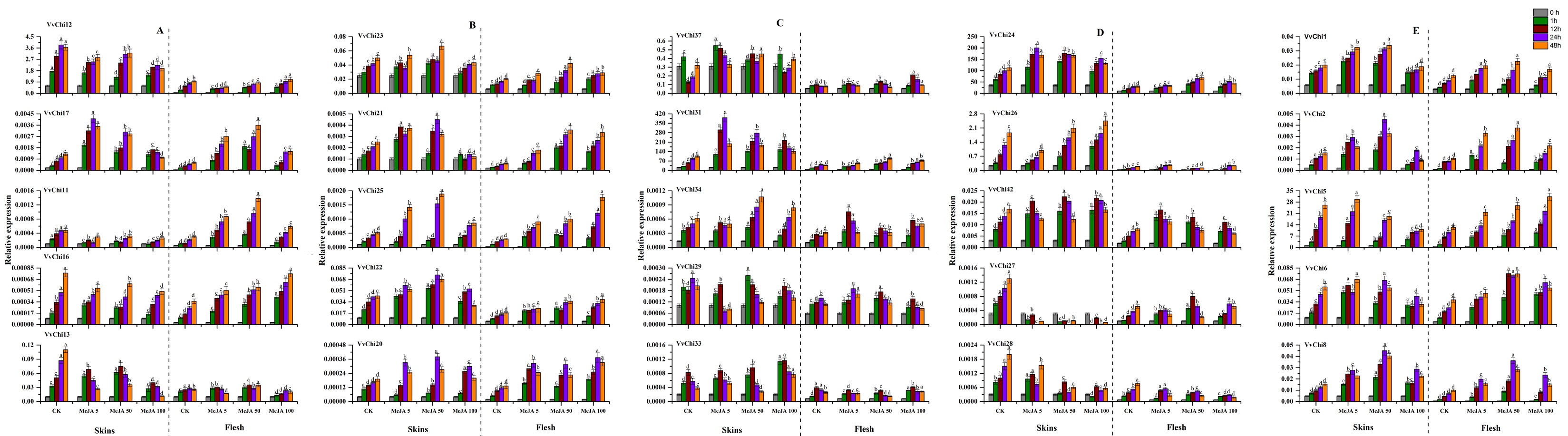
In view of the important effects of ETH and MeJA on grape growth and disease resistance, we studied the effects of ETH and MeJA treatments on the expression pattern of VvChis. The results proved that VvChis can respond to both hormones, showing different expression trends with the species and concentration of hormone (Figure S5, S6). Overall, VvChis is more sensitive to 500 mg·L− 1 ethylene and differs in skin and fruit. The family classification has a great influence on the expression of VvChis: GH18 has an up-regulation trend under both hormone treatment, and more sensitive to ETH 500 mg·L− 1 and MeJA 50 µmol·L− 1; GH19 members vary greatly, VvChis of group D were sensitive to ETH 500 mg·L− 1, while group C more sensitive to MeJA 5 µmol·L− 1, and others have no obvious features.
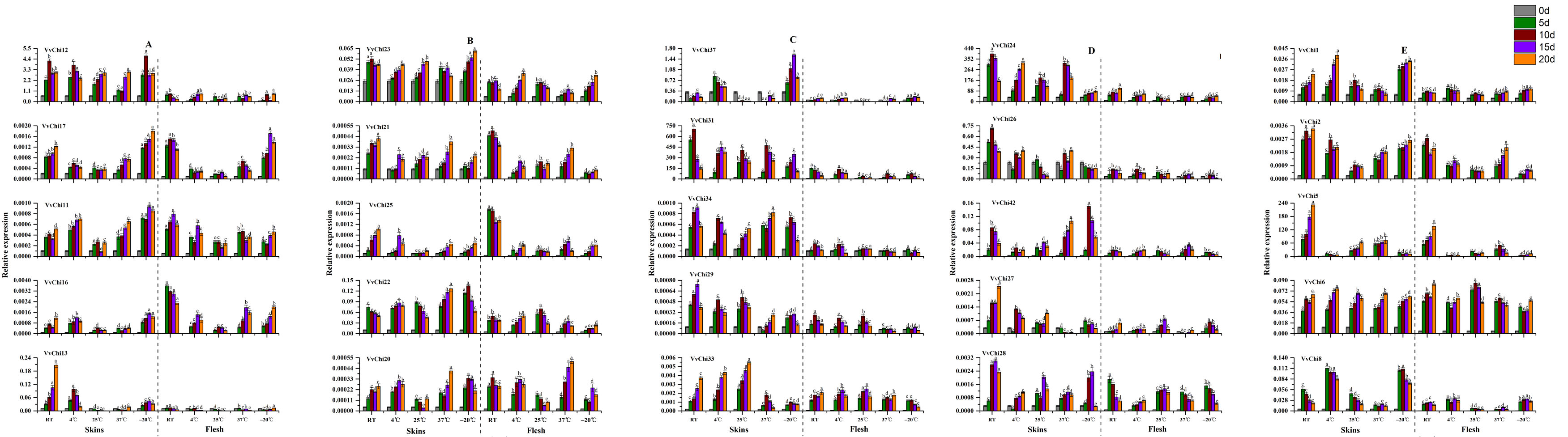
The induction of chitinase by low temperature is more sensitive than high temperature (Figure S7). The reaction of flesh and skin were not synchronized to the temperature, and flesh was more sensitive. The response to temperature was different between different groups: Group A was higher under high temperature; Group B caused significant up-regulation in skin at 37 °C and − 20 °C; Group C were up-regulated at 4 °C in skin; VvChis in group D were significantly affected by temperature; Group E were higher in skin at higher temperature except VvChi5. The enzyme activity assay showed that the temperature had significant effects on SOD, POD, CHI, APX and β-1,3 GA, which were generally higher at 37 °C. Different from other enzymes, CHI had the highest activity at -20 °C.
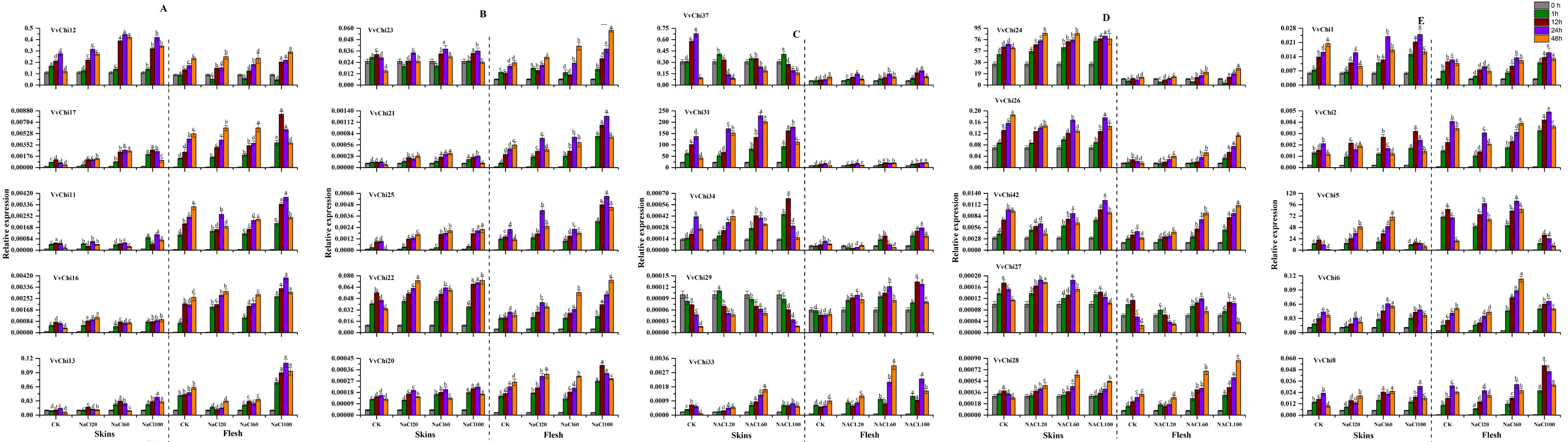
VvChis showed an up-regulation trend after treatment with different concentrations of NaCl, and more sensitive to high concentrations (Figure S8). On the other hand, NaCl treatment increased the activity of five enzymes during short time, and then slowly decreased, which had little effect on SOD and β-1, 3 GA, and obvious effects on POD, APX and CHI. Among these, CHI activity is consistent with POD, but different from β-1,3 GA.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}