Identifying correlative biomarkers with high sensitivity and specificity is of great importance in cancer diagnosis and treatment (5). Recently, with the development of next generation sequencing, lncRNAs are revealed as multifaceted regulators in a wide range of cellular homeostasis (37) via participating in epigenetic, transcriptional and posttranscriptional regulation of genes (26). Increasing evidences have established a strong relationship between dysfunction of lncRNAs and cell fate determination as well as disease pathogenesis, such as aging (38), arthritis (39), and cancer (13). LncRNA MIR31HG, locating at human chromosome 9p21.3 (40) with transcription regulated by methylation of the promoter region (27), was found aberrantly expressed in several types of cancers. Specifically, MIR31HG was found elevated expressed in breast cancer (13), cervical cancer (28), chordoma (16), osteosarcoma (15), lung cancer (2, 23, 41), OSCC (4), PDAC (7), VSCC (29) and LSCC (36, 42, 43). Nevertheless, the expression levels of MIR31HG was reported down-regulated in triple-negative breast cancer (TNBC) cell lines of basal subtype (27), bladder cancer (26), gastric cancer (25), CRC (32, 33), and HCC (6). Besides, abnormal expression of MIR31HG was deemed as potential biomarker to reflect the clinicopathological characteristics and prognostic outcomes of cancer patients (28). For instance, increased MIR31HG expression was found both in LUAD/ NSCLC tissues when compared with counterparts, and positively associated with unfavorable TNM stage, metastasis, histological differentiated degree, and forecasted shorter OS (2, 23, 41). Notably, MIR31HG expression was even higher in gefitinib-resistant NSCLC cells, and knockdown of MIR31HG could promote cell apoptosis and cell cycle arrest, and subsequently induce gefitinib sensitivity (20). More recently, He J et al. further verified that MIR31HG was also remarkably increased in gefitinib-resistant NSCLC patients, and MIR31HG overexpression contributed to reduced sensitivity of NSCLC cell to gefitinib in vitro (44). By contrast, Yan S et al. reported that overexpression of MIR31HG significantly inhibited HCC proliferation and metastasis in vitro and impeded tumorigenesis in vivo (6). In CRC, the MIR31HG expression was negatively correlated with unfavorable prognosis, as indicated by advanced pathologic stage, and lager tumor size (19). Consistently, Li Y et al. found that high expression of MIR31HG predict high OS and DFS in CRC patients treated with oxaliplatin (32).
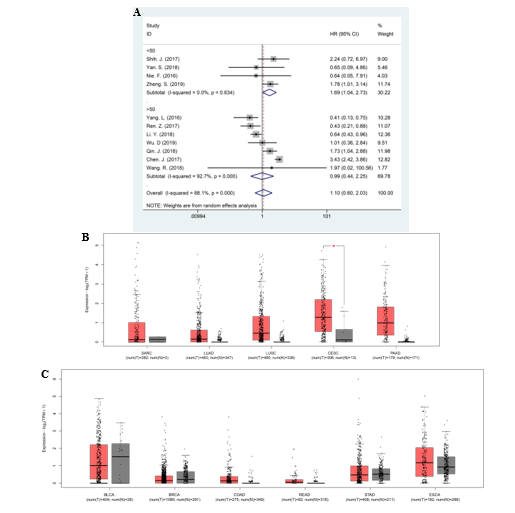
Given the prognostic significance of MIR31HG in cancers, in the current study, we firstly explored the association between MIR31HG level and clinical outcomes by performing a meta-analysis containing seventeen literatures with 2,573 patients. The pooled results of subgroup analysis as per the tumor types demonstrated that high MIR31HG expression predicted unfavorable OS in patients with lung cancer and other cancers, and poor RFS in all selected studies, respectively. On the contrary, overexpression of MIR31HG was closely associated with favorable OS and DFS in gastrointestinal cancer, indicating a tissue-specific predictive prognosis of MIR31HG in multiple human cancers. Besides, the relationship between expression of MIR31HG and other parameters were analyzed. Consistent with the predictive value of MIR31HG in OS, high level of MIR31HG was also significantly correlated with advanced clinical stage and LNM potential in lung cancer. Previously, the remarkable associations between MIR31HG expressions with LNM risk or tumor stage in gastrointestinal cancer were not noted. However, we found that the study performed by Sun. K. et al. may introduce bias to this meta-analysis, as shown in sensitivity analysis. Therefore, in order to increase the stability of the results, we excluded this study and found that elevated expression of MIR31HG were negatively associated with clinical stage and LNM risk in gastrointestinal cancer. However, no correlation between MIR31HG expression and other clinical characteristics, such as DM and tumor size, were noted. It is worth mentioning that since the studies with regard to the relationship between expression of MIR31HG and other clinical parameters are limited, these results could be biased and therefore needs further confirmation in future studies. Beside, in order to increase the credibility of the results, we performed cross-validation of the results by using TCGA dataset. The results showed the elevated MIR31HG was significantly correlated with poor OS rather than DFS in lung cancer. However, it should be noted that the enrolled studies reporting MIR31HG expression in lung cancers consisted of LUAD and NSCLC, while the cancer types in TCGA were LUAD and LUSC, which may lead to potential difference in predictive value of MIR31HG on clinical outcomes between this meta-analysis and TCGA dataset. Moreover, the limited sample size in TCGA may cause possible bias to the results.
Mechanistically, lncRNA could execute versatile functions in diseases through alternative splicing, epigenetic modulation, chromatin modification, scaffolding/decoy function or acting as molecular sponge (45). To begin with, given the critical role of MIR31HG in hypoxia-associated malignant progression, MIR31HG was observed to promote tumorigenesis by serving as a HIF-1α co-activator and regulating HIF-1 transcriptional network (4, 42). For instance, in LSCC, MIR31HG could facilitate cell proliferation, cell cycle progression and suppress apoptosis via HIF-1α and p21 (36). In OSCC, MIR31HG knockdown impaired the HIF-1α transactivation, sphere-forming ability, metabolic shift and metastatic cascade both in vitro and in vivo (4). Besides, lncRNA could involve in the competing endogenous RNA (ceRNA) regulatory work to function as endogenous miRNA sponge (Table 3) (6). As the host gene of miR-31, MIR31HG was firstly identified to co-express or modulate the expression of miR-31 in certain cancers (44). For example, MIR31HG could enhance proliferation, migration and invasion by up-regulating EZH2/miR-31 and then indirectly activating the oncogene RNF144B in chordoma (16). Yang S et al. demonstrated that elevated MIR31HG could inhibit miR-31 expression, and increase RhoA in LSCC, and via which promote cell growth, cell cycle and invasion (43). In addition to miR-31, other miRNAs were also identified to interact with MIR31HG in carcinogenesis. It was reported that MIR31HG could directly sponge to tumor suppressor- miR-361, rather than miR-31, and in turn regulate cell proliferation, cell cycle arrest, and apoptosis via targeting VEGF, FOXM1 and Twist in osteosarcoma (15). In NSCLC, MIR31HG behaved as an oncogene by inhibiting miR-214 expression, thereby facilitating cancer cell migration and invasion (41). In HCC cancer, MIR31HG could competitively bind miR-575, and positively regulate ST7L to impair cell proliferation and metastasis (6). Moreover, MIR31HG could regulate tumorigenesis by targeting a broad spectrum of target genes or pivotal signaling pathways (Table 3). In gastric cancer, MIR31HG may suppress cell proliferation and hinder tumorigenesis partly by regulation of E2F1 and p21 expression (25). In NSCLC, MIR31HG was found to enhance Wnt/β-catenin pathway, and induce epithelial-mesenchymal transition (EMT) phenotype (2). Jin C et al. identified that MIR31HG was induced by nuclear translocation of NF-κB, and in turn directly bind to IκBα and participated in NF-κB activation, revealing an interaction between MIR31HG and NF-κB in osteogenic differentiation (46). Of note, MIR31HG could modulate gefitinib resistance partly through EGFR/PI3K/Akt (20, 44) or RAF-MEK-ERK pathways (44). Furthermore, Zheng S and colleagues discovered that down-regulation of MIR31HG inhibited NSCLC cell proliferation, invasion and EMT phenotype via up-regulating E-cadherin expression, inhibiting Wnt/β-catenin cascade, and down-regulating expression of Twist1 and vimentin (2).
Despite an effort to make a comprehensive analysis, limitations remained inevitably in this study. First, most of the studies were among Asian populations though we do not impose restrictions on region during literature retrieval, which may introduce possible geographical bias. However, since these results have been further confirmed by TCGA dataset (21), it could be more credible to generalize the data to other populations. Second, some HRs with corresponding 95% CIs were indirectly extracted from the Kaplan–Meier (K-M) survival curves, which may introduce possible bias. Third, the cut-off value of MIR31HG levels varied among the selected studies, which may contribute to potential limitation to the clinical use.
{kind=link}