The possibility of combining non-food production and phytoremediation arouses more and more interest. The implementation of this strategy requires the selection of fast-growing crops that can accumulate and tolerate HM. Hemp (Cannabis sativa L.) is a multi-uses plants providing cortical fibres used in the manufacture of various products, and hurds that enter in the composition of building materials (Deleuran and Flengmark 2006), and is thought to be able to reduce HM contamination in soils thus contributing to the remediation of contaminated sites (Angelova et al. 2004; Bona et al. 2007; Shi et al. 2009 and 2012; Ahmad et al. 2016; Kumar et al. 2017).
The present study confirmed that Zn significantly accumulated in C. sativa cultivated in nutrient solution under Zn excess. However, growth properties were only marginally affected by Zn excess: the recorded reduction in fresh and dry weight remained not significant from a statistical point of view, considering the registered variability between plants but was clearly detectable (see Fig. 1). It has however to be mentioned that we used a short-term exposure of one week and it can therefore not be excluded that a longer duration would lead to a significant yield decrease.
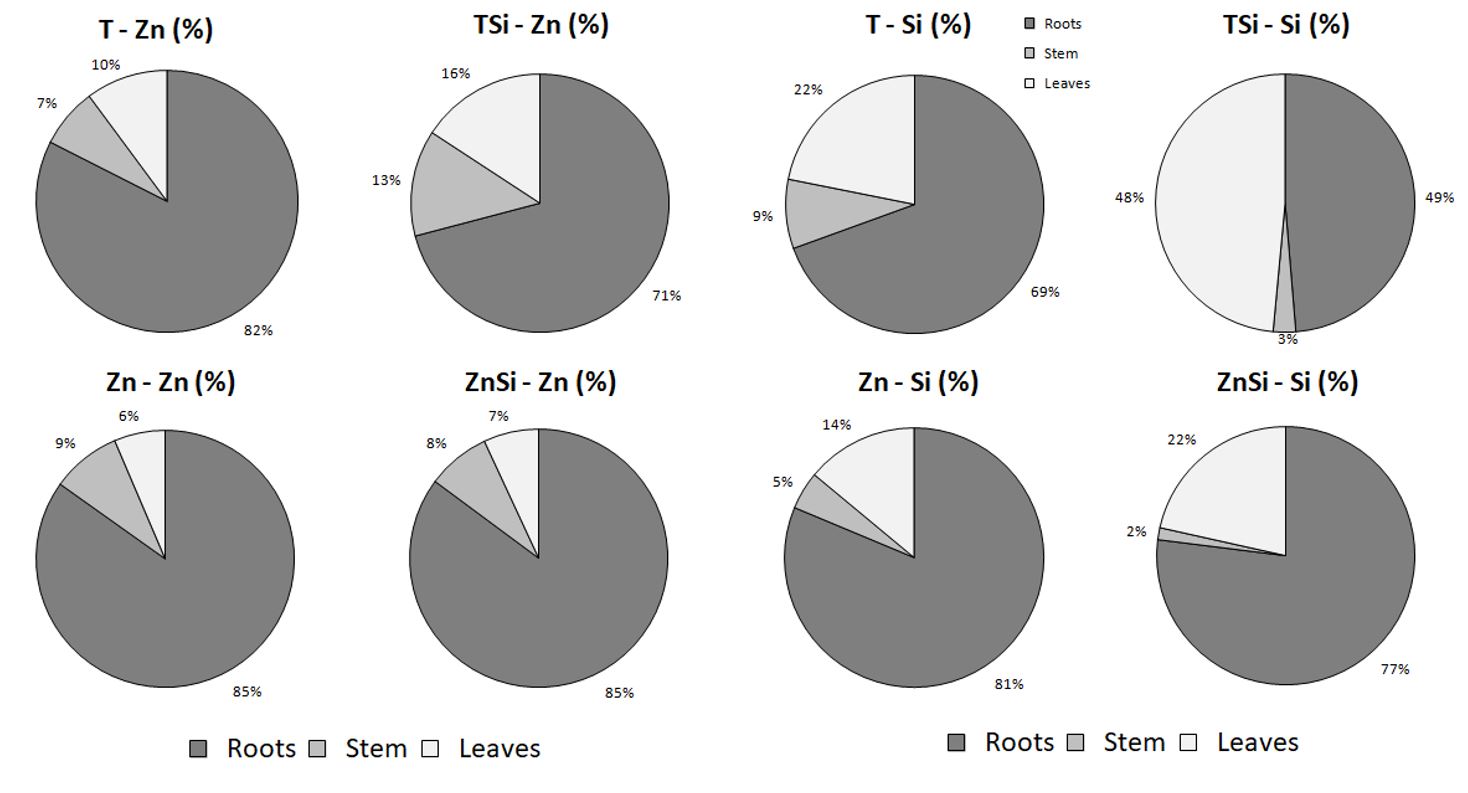
In plants exposed to Zn excess, Zn concentration in stem was higher than in the leaves. This could be linked to the fixing properties of the fibres which are exploited for biosorption purposes by numerous authors (Pejic et al. 2011; Vukcevic et al. 2014a, b). Zinc is indeed able to bind to carboxyl and hydroxyl groups of cell wall polymers such as those occurring in bast fibres and Loiacono et al. (2018) recently demonstrated that hemp fibres may be efficiently used to clean up waste water contaminated by numerous HM. Wall retention could also occur in planta, during the formation of the fibres, although this implies that HM need to be in close contact with the fibre and bast fibres are located in the phloem rather than in the xylem where transpiration stream occurs. More accurate techniques of histological localization would allow us to precise the location of Zn in situ.
Zn translocation from roots to shoots, estimated on a concentration basis (TFc), was however limited (TFc ~ 0.1) but it could also be interesting to estimate TF on a total amount basis (TFa, Table 3) based on the quantities actually exported from the substrate (Ali et al. 2013). For all treatments, translocation factor estimated on a total amount basis (TFa) was higher than translocation factor estimated on a concentration basis (TFc) but remained always lower than 1. This suggests that, despite a good level of tolerance, hemp adopted an excluding strategy. A similar observation has already been reported by Angelova et al. (2004), Löser et al. (2002) and Shi et al. (2009) in hemp exposed to HM. Root sequestration is a strategy widely developed by non-hyperaccumulating plants to avoid the accumulation of toxic elements in photosynthetic tissues. Hence, the use of Cannabis sativa cv Santhica 27 in phytoremediation should be restricted to phytostabilization but not phytoextraction of Zn from contaminated soils. This implies that hemp is also able to display tolerance mechanisms.
We demonstrated that metallic stress had only a slight negative impact on biomass production (FW, DW, total leaf number, stem length) and pigment concentration, although we did not observe any mortality in plants nor a significant impact on photosynthesis-related parameters. According to Piotrowska-Cyplik and Czarnecki (2003) and Linger et al. (2005) a reduction of pigments content, and therefore in the availability of photoassimilats, as well as the energy cost of tolerance mechanisms, may explain the reduced biomass production (Ghnaya et al., 2009). According to Küpper et al. (1998), Zn may substitute to Mg within chlorophyll and this could explain both an alteration in the efficiency of light phase, and a decrease in unstable chlorophyll prone to degradation by chlorophyllase. This, however, is considered to mainly occur in response to Mg deficiency while we demonstrated in the present study that Zn excess reduced the Mg content in the leaves by only 5.2% (Table 3). Accordingly, data provided by chlorophyll fluorescence indicated that the light phase by itself is only marginally affect by Zn excess.
We also paid attention to plant water and antioxidative status. Water content was not affected by the treatments applied but Zn excess significantly lowered ΨS values in leaves, which is a clear indication that adaptations were required for Zn-treated plants to maintain internal water potential and turgor. Such accumulation of osmotic compounds contributes to turgor maintenance and CO₂ diffusion within the mesophyll. This probably explain stable Ci values in plants exposed to Zn excess compare to controls while other gas exchange parameters tended to decrease. Beside CO2 diffusion within the mesophyll, the highest resistance encountered by CO2 to reach the sites of carboxylation is located at the stomatal level. Zinc tended to decrease the gs value but this had no impact on mean Ci which suggest that the carboxylation efficiency might be hampered by Zn excess. Indeed, Zn-treated plants exhibited the lowest A value: Cambrollé et al. (2013) mentioned that Zn may also substitute to Mg in Rubisco itself, leading to under-utilization of ATP and NADPH in CO2 fixation. Nevertheless, this hypothesis might be invalid in our case since no important Mg decrease was recorded in the leaves. Data are still missing regarding the impact of Zn on activity of other enzymes involved in the Calvin cycle.
The present study also showed that hemp exposed to Zn excess encountered a moderate oxidative stress: MDA content in leaves remained similar to controls, but increased in roots. Zinc accumulated to higher amounts in the roots than in the leaves; hence, a higher oxidative stress in the below part of the plant was not unexpected. This suggests that, in leaves, oxygen species were quickly scavenged. Reduced glutathione (GSH) is involved in the reduction of an important part of ROS generated due to stress (Shao et al. 2007). In our study, HM exposure led to higher GSHt concentration in leaves and elevated GSSG/GSH ratio, suggesting that GSH helped to withstand oxidative stress in leaves. This may also explain the decrease of total antioxidant capacity, GSH being part of the AOAM fraction. Besides its role as an antioxidant, GSH acts as a precursor of phytochelatin (PC) synthesis. In our study, HM exposure led to higher PC content in leaves. Chelating metals by forming PCs or metallothioneins (MTs) metal complex at the intra- and intercellular level are part of the mechanisms used by plants to counteract HM toxicity (Alloway et al., 2013; Citterio et al., 2003; Tennstedt et al. 2009; Emamverdian et al., 2015). Nevertheless, a higher PC concentration in the leaves than in the roots is rare in plants exposed to HM. Wogkaew et al. (2019) suggested that glutathione may be involved in the translocation of Zn from the root to the shoot: the underlying mechanisms however, remains unknown and this observation did not explain why, in the present case, Zn remained sequestered in the roots while glutathione accumulated in the leaves. Besides, oversynthesis of glutathione requires large quantities of S. Hemp increased the S accumulation in roots, stems and leaves in the presence of Zn excess confirming that hemp is able to trigger this protective mechanism. However, it is important to keep in mind that to combine phytostabilization with non-food production, the properties of the harvested biomass have to remain compatible with the requirements of industry from a qualitative point of view (Luyckx et al., 2019). Additional work is therefore needed to precise the impact of Zn excess on valuable plant parts.
Although Si is not considered as essential for plants, it intervenes as a beneficial element in their defence and growth. As such, it is used in agriculture as a biostimulant. Hemp is not considered as an Si accumulator under normal conditions. However, in this study, Si significantly increased root and leaf Si concentration in the absence and in the presence of Zn excess. In roots, plants of CSi treatment accumulated 3.6x more Si than plants of C treatment while ZnSi treated plants accumulated 7.4x more Si than plants of Zn treatment. This suggests that plants stimulated Si uptake to cope with Zn stress. In rice exposed to heavy metals and Si, Kim et al. (2017) and Ma et al. (2015) reported an increase in the expression of genes involved in the transport of Si (OsLSi1 and OsLSi2) to improve resistance to metal stress. In the present case, however, high Si accumulation in the root of ZnSi-treated plants did not protect the roots from oxidative stress since MDA concentration was similar to Zn-treated plants.
H2SiO3 application under Zn exposure interfered with HM absorption. Zn concentration was clearly lower in roots, stems and leaves of Si-treated plants in the presence of Zn excess than in plants exposed to Zn in the absence of Si. A first hypothesis is that the presence of Si in the nutrient solution reduces Zn availability through the precipitation of Zn silicate. Bokor et al. (2014) indeed mentioned that Zn2SiO4 may occur in water experiments but this is not confirmed by the speciation program VISUAL MinTEQ which clearly indicated that, for the range of concentration and pH of the solution used in our study, Zn remained fully soluble in the solution. The decrease of Zn concentration cannot be attributed to a decrease in transpiration rate since no significant difference for E values was observed between ZnSi and Zn treatments. It has already been observed in rice plants treated with Cu/Cd a decrease in the expression of HM transporters in the presence of Si (Kim et al. 2017; Ma et al. 2015). Huang and Ma (2020) recently demonstrated in rice that Si supply decreased Zn concentration in both the root and the shoots: according to these authors Si acts on Zn uptake by down-regulating OsZIP1 implicated in Zn uptake. The comparison should however be established with caution considering that rice is a specific plant species for Si hyperaccumulation and that data obtained with rice are not necessarily valid for other plant species, especially dicots. Beside Zn absorption, some authors reported that Si may reduce Zn translocation from the root to the shoot (Zajaczkowska et al. 2020; Naeem et al. 2015) but this was not observed in our experiment since TF value for Zn were hardly modified by Si. This implies that as far as hemp is concerned, Si may impact transporters involved in Zn uptake (especially those encoded by ZIP genes) but had no impact on transporters involved in Zn xylem loading and long-distance transport (HMA2 and HMA4) (Zlobin 2021).
It is also interesting to notice that the recorded Zn-induced increase of Zn accumulation was higher than the Si-induced decrease of Zn accumulation in plants of ZnSi treatment. Hence, if Si contributes to defence and growth, silicon could be compatible with phytostabilization purposes. Zn accumulation in control plants was not significantly affected by Si. Although no significant difference was recorded following H2SiO3 application, Si slightly improved gas exchange (gs, A, E) in controls and net photosynthesis under Zn exposure. In the same treatment (ZnSi), the fact that NPQ was always higher than in Zn treatment may indicate that plants quickly dissipate an excess of energy in order to maintain an adequate balance between photosynthetic electron transport and carbon metabolism (Gharbi et al., 2017). Si paradoxically decreased pigments content in control plants which was an unexpected result.
Exogenous application of Si under Zn excess increased total antioxidant capacity (AOAM and AOAD fractions) and decreased MDA content mainly in the leaves, as already observed by Kim et al. (2017). This suggests that Si may trigger oxidative tolerance processes in hemp. The same beneficial effect of Si was noticed in leaves of control plants. Increased antioxidant capacity under Si exposure suggested a higher GSHt content. However, in the present study, under Zn excess GSHt content decreased following Si application. The increase in the antioxidant capacity despite a decrease in GSHt content may be due to activation of other antioxidants by Si: the fact that both AOAD and AOAM antioxidant activities increased in response to Si suggests indeed that other compounds, such as ascorbate and α-tocopherol, which were not quantified in the present study, may increase in response to exogenous Si. Moreover, the lower Zn accumulation induced by Si probably decreased ROS production and the need of GSH as an antioxidant and a precursor of PC synthesis. PC content in leaves was indeed decreased in plants of ZnSi treatment comparatively to Zn–treated plants, while we observed an opposite trend in roots, although Si decreased Zn accumulation in both organs. This suggests that PC more efficiently sequester Zn in the leaves than in the roots: while Zn concentrations in the roots are quite high, PC contents were unexpectedly low. Using K-edge extended X-ray absorption fine structure (EXAFS) spectroscopy measurements, Lefèvre et al. (2016) demonstrated that in the roots of HM resistant species Zygophyllum fabago, Zn mainly coordinate to Zn–O/N–C groups suggesting that PC did not play a key role in Zn tolerance in this organ and that other compounds, such as organic acid or polyamines, may be involved.
As a matter of fact, exogenous Si provides some metabolic advantages to hemp exposed to Zn excess, especially in relation to a decrease in Zn accumulation in the different parts of the plant. However, this was not sufficient to significantly increase plant growth on a short-term basis. Bokor et al. (2014) found similar data regarding specific maize cultivars simultaneously exposed to Zn and Si. According to these authors, increasing concentration of Si in combination with Zn treatment even increased physiological stress in comparison to Zn treatment. Similar results were observed by Masarovic et al. (2012) in sorghum who observed no positive effect of Si on high Zn in the medium.
{kind=link}