The high density of TANs was related to the poor prognosis of breast cancer patients.
To investigate the presence of TANs in breast cancer, clinical breast cancer surgical samples (table S1) were obtained from the two institutions and IHC was performed for CD66b. CD66b+ neutrophils infiltrating in the parenchyma of breast cancer tissue counted (Fig. 1A). Median value of the density of TANs was chosen to be the cut-off value to divided into 2 groups.
The prognostic value was then analyzed by survival and Cox multivariate analysis. In all 3 cohorts, high density of TANs was positively correlated to poor PFS for BC patients (training cohort: Fig. 1B, P < 0.0001; validation cohort: Fig. 1E, P < 0.0001; independent cohort: Fig. 1H, P = 0.0221 < 0.05). The univariate analysis showed significant positive association between TANs and PFS in all three cohorts (Table 1). Cox multivariate analysis found that high density of TANs was an independent prognostic factor for poor PFS in all 3 cohorts (training cohort: Fig. 1D, Hazard ratio (HR) = 2.8, P < 0.001; validation cohort: Fig. 1G, HR = 4.0, P < 0.001; independent cohort: Fig. 1G, HR = 3.5, P < 0.001).
The prognostic value TANs were then assessed by ROC analysis. TANs showed a high prognostic value in predicting poor PFS in all training [Fig. 1C, Area under the curve (AUC) = 0.80, 95% confidence interval (CI): 0.74–0.87)], validation (Fig. 1I, AUC = 0.72, 95% CI: 0.75–0.90) and independent (Fig. 1F, AUC = 0.82, 95% CI: 0.61–0.82) cohorts. The prognostic value of TANs for overall survival were then validation in TCGA datasets. According to the TIMER, high density of TANs was positively correlated to poor OS (Fig. 1K, P = 0.018, HR = 1.36).
To further confirm our findings, 1222 breast cancer patients RNA-seqs from Metabric were downloaded and analyzed. High level of TANs positively related to poor PFS (Fig. 1G, P = 0.023 < 0.05). The univariate analysis found high level of TANs to be associated with worse PFS (P = 0.023, HR = 1.29, Supplementary table S2). Cox multivariate analysis also demonstrated that high level of neutrophils was an independent prognostic factor (Fig. 1M, P = 0.018, HR = 1.3).
Activated by BC cells, TANs promoted MCF-7 activities
Neutrophils were isolated from peripheral blood from healthy donates as reported previously[13]. The purity of neutrophils isolated was more than 90% (Fig. S1). Compared with unstimulated neutrophils, a significant increase in the lifespan of neutrophils treated with either MCF7CS (MCF7TANs, P = 0.0111) or MDA231CS (MDA231TANs, P = 0.0003) was evident. However, the lifespan of MDA231-TANs was significantly higher than that of MCF7TANs (Fig. 2A).
We further evaluated the role of TANs on breast cancer cells. After culturing breast cancer cells with supernatant from either MDA231TANCS and or MCF7TANCS, there was no significantly change was found in the migration and invasion of both MDA-MB-231 and MCF-7 cells respectively (Fig S2). However, MDA231TANCS was found to promote the proliferation (Fig. 2B, P < 0.0001), migration (Fig. 2C, P = 0.0043) and invasion (Fig. 2D, P = 0.0011) abilities of MCF-7 cells (Fig. 2B, 2C, 2D), hence MDA231TANCS was chosen for further investigations.
Neutrophils were activated by tumor-derived G-CSF and turned to TANs in breast cancer
Based on the above findings, the effective components of MDA231CS were then investigated by cytokine antibody arrays. Compared to MDA231CS, the level of G-CSF, CXCL1, GDF-15, and PDGF-AA was significantly lower in TANCS (Fig. 3A).
With all 4 solute mediators, both viability and pro-invasive abilities of TANs were promoted. To identify the most key component, neutrophils were then stimulated by 3 of these 4 cytokines in turn. As results suggested, without rh-G-CSF stimulation, both viability and migration- promotion ability of neutrophils will not be promoted (Fig. 3B, 3C, Fig S3, Fig S4). To further investigate the role of G-CSF, neutrophils were then stimulated by rh-G-CSF. As the result indicated, its viability and abilities of pro-invasive and pro-metastasis were significantly promoted. (Fig. 3B, 3C).
To confirm the role of G-CSF being the main component in activating neutrophils, G-CSF vector was transfected into MCF-7 cells (GCSFhigh- MCF7) to up-regulate its expression of G-CSF. After the exposure to GCSFhigh- MCF7 supernatant, the viability of neutrophils was significantly enhanced. Significantly increased invasion and migration of MCF-7 cells were then observed after the cancer cells were cultured in the supernatant of GCSFhigh- MCF7- TANs (Fig. 3D, 3E).
The relationships between G-CSF and TANs were then investigated in breast cancer surgical tissue. 20 fresh breast cancer tissues and their FFPS tissue were collected. The level of G-CSF was investigated by ELISA, and the density of TANs was evaluated by IHC. The results showed a positive correlation between these two factors (Fig. 3F, P = 0.012, rs=0.553), which was also demonstrated in breast cancer RNA-seq from TCGA (Fig. 3G, P < 0.0001, rs=0.156).
PI3K-AKT and NFƙB signaling pathway were activated in TANs by G-CSF
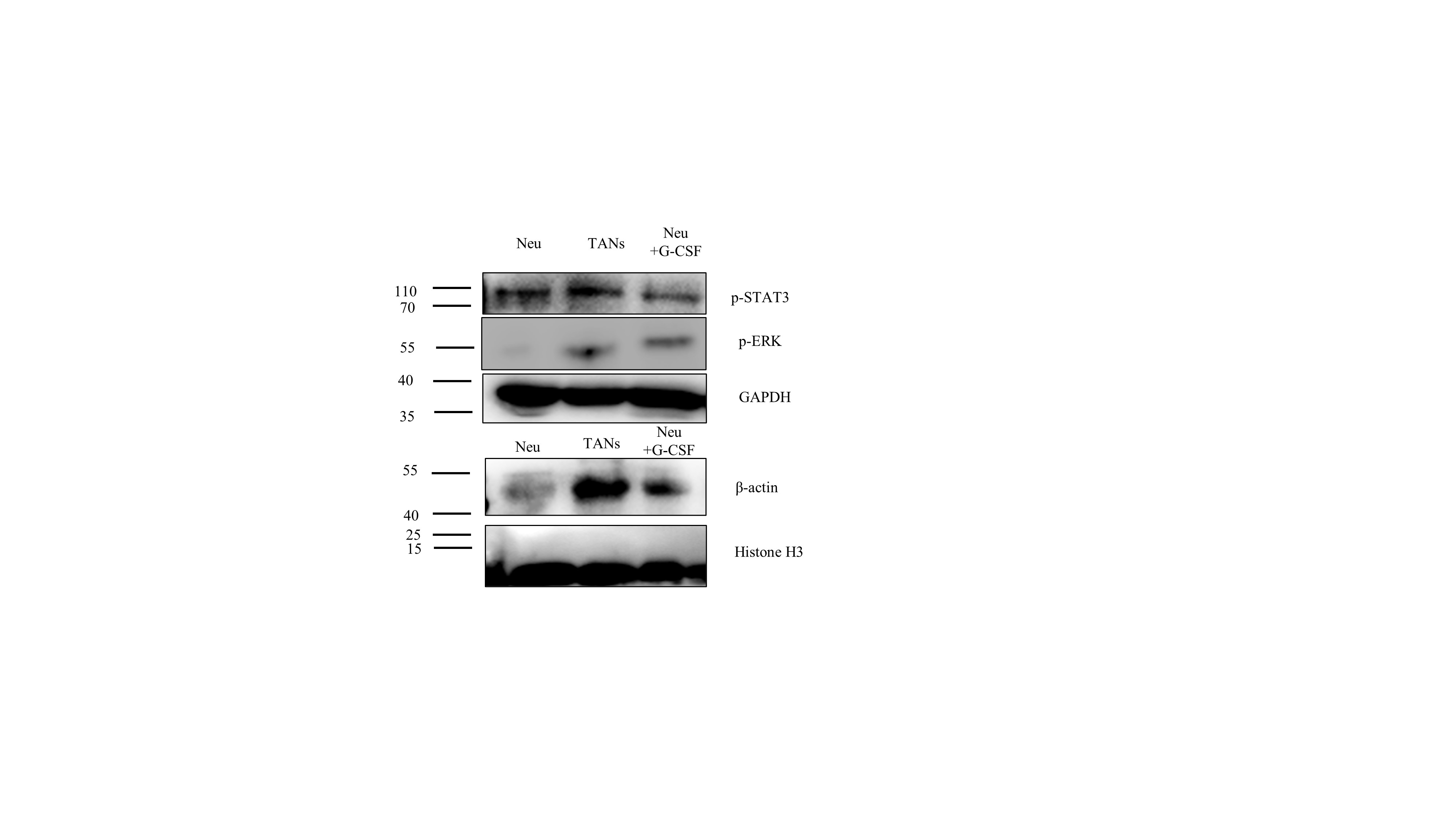
The underlying mechanism mediating G-CSF action on neutrophil activation was investigated. Several signaling pathways such as PI3K- AKT, JAK- STAT3, MAPK - ERK1, NFκB and Wnt- β-catenin have been shown to regulate the action of G-CSF on the activation of neutrophils. [14–18]. Compared with neutrophils, levels of PI3K, p-AKT and nuclei- NFκB in TANs and GCSF-TANs were significantly increased. (Fig. 4A, Fig. S5).
After exposure to the MK2206 (AKT inhibitor), the lifespan of TANs was significantly decrease, while the ability to promote tumor activities was not affected. In contrast, after exposing to the JSH-23 (NFκB inhibitor), the ability to promote tumor activities of TANs was a significant decrease, while the lifespan was not affected. Exposure to both inhibitors, neutrophils cannot be activated to TANs by both MDA231CS and rh-G-CSF (Fig. 4B, 4C).
The roles of PI3K-AKT and NFκB were further confirmed in female breast ca ncer tissues by using IF assay. The results showed positive staining signals of p-AKT (Fig. 4D) and nuclei-NFκB (Fig. 4E) located in CD66b + neutrophils, which infiltrated in the parenchyma of breast cancer tissue.
TANs promoted the migration and invasion of MCF-7 cells by activating the RLN2-PI3K-AKT-MMP-9 pathway
The role of TANs on breast cancer cells were then investigated. According to the cytokine array assay, there was a significantly increased expression levels of IL-1ra, M-CSF, MIF, RLN2, VEGF and sCD31 in TANCS (Fig. 3A), which were confirmed by ELISA (Fig. 5A).
After stimulated by all these 6 cytokines together with NeuCS, the migration and invasion of MCF-7 were promoted. To identify key components, MCF-7 cells were then stimulated by NeuCS with 5 of these 6 cytokines in turn. As the results showed, without RLN2 stimulation, the invasion and migration of MCF-7 cells cannot be promoted (Fig. 5B, S6). To identify the role of RLN2 in TANCS, MCF-7 cells were then treated with NeuCS + RLN2. Transwell assay showed, both of the migration and invasion of MCF-7 cells were significantly promoted, while treated with rh-RLN2 alone, no promoted migration and invasion were observed (Fig. 6A). As it had been known that RLN2 targeted on RXFP1 on breast cancer cells, its expression in MCF7 were then knocked down by (siR-MCF7). The MCF-7 cells transfected by si-NC was taken as control (siN-MCF7, Fig. 6S). Treated with TANCS and NeuCS + rh-RLN2 respectively, the migration and invasive of siR-MCF7 cells cannot be promoted, while these two supernatants still promoted this behavior for siN-MCF7 (Fig. 6B).
As it had been known that the RLN2 promoted the breast cancer cell migration and invasive by activating the PI3K-AKT-MMP9 signaling pathway [19], the role of RLN2 pathway were studied. Levels of expression for PI3K, p-AKT and MMP9 in siN-MCF7, TAN-siN-MCF7, Neu-siN-MCF7, Neu + rh-RLN2-siN-MCF7 and TAN-siR-MCF7 were evaluated by WB. As the results showed, levels of PI3K, p-AKT and MMP9, compared to the siN-MCF7 and Neu-siN-MCF7, were significantly increased in TAN-siN-MCF7, Neu + rRLN2hR-siN-MCF7 respectively. Significantly lower levels of PI3K, p-AKT and MMP9 were found in TAN-siR-MCF7 compared to those in TAN-siN-MCF7. After inhibiting AKT in TAN-siN-MCF7 by MK2206, the level of MMP-9 was significantly decreased (Fig. 7A). After knocking down MMP9 in MCF7 cells by siRNA (siM-MCF7), the migration and invasive of the cancer cells were not promoted, regardless of the treatment of TANCS or NeuCS + RLN2 (Fig. 7B).
The relationships of TANs, RLN2 and MMP9 were evaluated in 20 breast cancer surgical tissues by IHC and ELISA. As the result showed, the density of TANs was positively related to the level the expression of RLN2 (rs= 0.3976, 95%CI: 0.0029–0.6851, P = 0.0490) and MMP9 (rs =0.4437, 95%CI: 0.0589–0.7137, P = 0.0263). (Fig. 7C).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}