Herbicides are heavily used in agricultural areas to control unwanted plant growth and subsequently increase crop yields. Atrazine (ATR; 2-chloro-4-ethylamino-6-isopropylamino-1,3,5-triazine) is a chlorotriazine herbicide and is one of the most heavily applied herbicides in the United States (U.S.) with over 32 million kg being applied annually (EPA, 2020). ATR is typically applied to corn, sorghum and sugarcane crops to control the growth of broad leaf weeds. After being applied to agricultural areas, ATR can enter local streams and rivers through run-off, ground water seepage, regional transport (evaporation) and may be deposited by rain water (LeBlanc et al., 1997; Tong and Chen, 2002). Although the U.S. Environmental Protection Agency (EPA) sets a limit of 15 µg/L (ppb) ATR (60-day average) and an in-field downwind buffer of 4.6 m from aquatic environments, ATR has been detected at levels above 300 ppb in some U.S. rivers (EPA, 2014; Belanger et al., 2016; EPA, 2020). These elevated concentrations have also been shown to last for several weeks during the spring season (EPA, 2014). Exposure to high concentrations of ATR is also compounded by the fact that the half-life of ATR ranges from six days to several months or years (Comber, 1999). ATR has also been detected for up to 22 years following its application due to its potential for soil sorption and high soil mobility (Walther, 2003; Jablonowski et al., 2011). This is of concern because ATR exposure can then have long-term impacts on non-target species, like aquatic organisms, for years.
Understanding the impacts of ATR exposure on non-target aquatic organisms has been the focus of many reviews (Giddings, 2005; Solomon et al., 2008; Van Der Kraak et al., 2014; de Albuquerque et al., 2020). Because ATR comes into contact with a broad range of aquatic organisms, it can cause sublethal effects to nontarget species and interfere with the behavior, morphology and physiology of many species at environmentally relevant concentrations. ATR has been shown to affect reproductive physiology and behavior. For example, acute exposure to ATR led to demasculinization and feminization of male frogs (Xenopus laevis), feminization of mussels (Elliptio complanata), altered sperm quality in zebrafish (Danio rerio) and an inability to detect and localize mate odors in crayfish (Faxonius rusticus) (Tavera-Mendoza et al., 2002; Hayes et al., 2003; Flynn et al., 2013; Belanger et al., 2017a; Bautista et al., 2018). Gunkel and Streit (1980) suggest that ATR enters the body of most aquatic organisms via the gills and can be concentrated in various organs and tissues. ATR has been shown to accumulate in the liver/hepatopancreas, gallbladder and ovaries in fish (Danio rerio, Tilapia sparrmanii, Coregonus fera, Cyprinus carpio) and in the visceral mass, foot and mantle of bivalves (Ancylus fluviatilis, Anodontites trapesialis and Corbicula fluminea) following an acute exposure (Gunkel and Streit, 1980; du Preez and Van Vuren, 1992; Jacomini et al., 2006; Xing et al., 2012; Al-Sawafi and Yan, 2013). ATR was also shown to cause DNA damage in cells of tissues where it is known to accumulate. Exposure to ATR causes biochemical changes and DNA damage in the cells of the hepatopancreas of D. rerio and liver of Prochilodus lineatus (Zhu et al., 2011b; Santos and Martinez, 2012). DNA damage was also detected in peripherally located antennule cells of crayfish (Faxonius virilis) following acute ATR exposures (Abdulelah et al., 2020). Because of where they are located, these appendages and cells presumably come in contact with ATR readily in their environment, leading to damage and long-term chemosensory deficits (Belanger et al., 2016). Changes in morphology, in addition to the expression and activity of detoxification enzymes and antioxidant biomarkers of the hepatopancreas (digestive gland) of crayfish (F. virilis, Procambarus fallax f. virginalis, Cherax destructor), were also noted following exposure to ATR and its metabolites (Velisek et al., 2017; Stara et al., 2018; Steele et al., 2018; Awali et al., 2019). Many invertebrates, including crayfish, rely heavily on the hepatopancreas for detoxification of xenobiotics following exposure.
The hepatopancreas of the crayfish serves as the main energy reserve for growth and molting. Additionally, it is the main site of digestion, absorption and storage of nutrients. It is also the main organ of detoxification of xenobiotic compounds (Holdich, 2002). Given that the central function of the hepatopancreas in crayfish is the digestion of food and protection of the body from toxins, the physical, physiological and morphological state of this organ can provide insights into the effects of xenobiotics (for review see Belanger et al. (2017b)). Crayfish hepatopancreatic tissue can be used to monitor the health of the crayfish and can indicate when the animal has been exposed to harmful substances (Xiao et al., 2014; Velisek et al., 2017). Following exposure to xenobiotics, the hepatopancreas bioaccumulates by storing them in intracellular vacuoles (Icely and Nott, 1992). Changes in the histological organization of the crayfish hepatopancreas may be used as a bioindicator of contamination (Popescu-Marinescu et al., 1997; Desouky et al., 2013; Koutnik et al., 2014; Stara et al., 2016; Belanger et al., 2017b; Stara et al., 2018; Laurenz et al., 2020). The hepatopancreas is formed by numerous tubules separated by connective tissues with four types of epithelial cells, including blister-like secretory cells (B cell) which form large vacuoles that channel off harmful substances (Abd El-Atti et al., 2019). Further, these large vacuolar structures have been recognized as secondary lysosomes that are involved in enzymatic breakdown of xenobiotics and in cellular autophagy (Brown, 1982). Given the importance of the hepatopancreas in detoxification, its morphology and physiology can be used to identify areas of the aquatic environment that are under the stress of pollution.
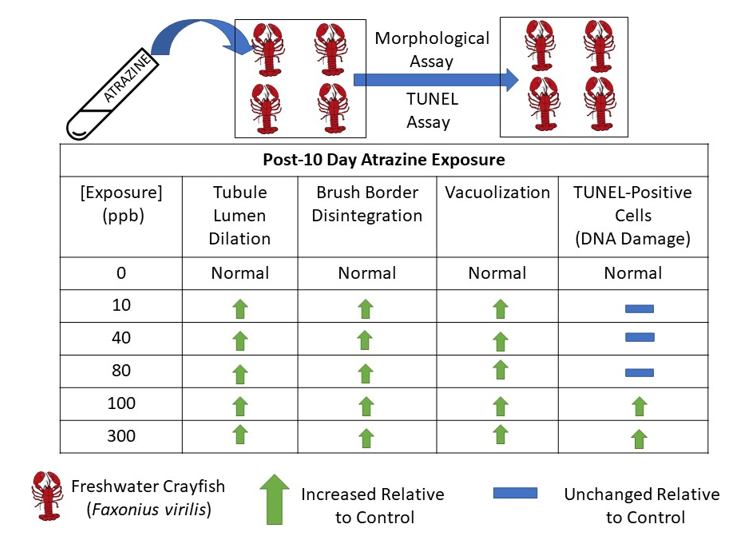
In this study, we investigated whether exposure to environmentally relevant concentrations of ATR (0, 10, 40, 80, 100 and 300 ppb) for 10 days caused morphological and cellular changes in hepatopancreas of crayfish. We hypothesized that as the exposure concentration increased, there would be changes in the morphology of the lobules of the hepatopancreas. Our hypothesis is based on studies indicating deterioration of the tubular epithelium and vacuolization of the hepatopancreatic lobules following exposure to the herbicides metolachlor and prometryne, and to pyrethroid insecticides (Wei and Yang, 2015; Stara et al., 2018; Stara et al., 2019). Moreover, because ATR exposure was shown to cause significant DNA damage, we also expected to see dose-dependent increases in hepatopancreatic cells with DNA damage following exposure to ATR (Liu et al., 2006; de Campos-Ventura et al., 2008; Cavas, 2011; Zhu et al., 2011b; Santos and Martinez, 2012; Abdulelah et al., 2020). To investigate DNA damage, we used a TdT mediated dUTP nick-end labeling (TUNEL) assay. In support of our hypothesis, we expected the percentage of TUNEL-positive cells in the hepatopancreas to increase in a dose-dependent manner. Additionally, we also expected to see an increase in vacuolization, tubular destruction and a decrease of microvilli in the hepatopancreas with increasing detoxification of ATR. Overall, it is important to understand how ATR-induced changes affect the morphology of the hepatopancreas, as overall physiological function is correlated with physiological condition (Popescu-Marinescu et al., 1997). Acute ATR exposure could lead to long-term changes in the crayfish’s ability to detoxify xenobiotics and subsequently affect survival.
{kind=link}