Effectiveness of the Pib gene and frequency of AVR-Pib alleles
The efficacy of the Pib gene was examined by pathogenicity assays. A total of 223 of the 366 M. oryzae isolates (4000 isolates were collected from six rice growing regions from 1997 to 2012 in Yunnan Province, and total of 366 isolates are selected from six rice growing regions as representative isolates.) tested were avirulent to the Pib gene-containing rice monogenic line IRBLb-B (Table 1). The percentage of avirulent isolates to Pib was 60.9%, whereas the remaining 143 isolates were virulent to the Pib gene (Table 1). The percentage of avirulent isolate was 100, 75.9, 75.0, 57.6, 53.6, and 48.2% in northwestern, central, northeastern, southeastern, southwestern, western Yunnan Province, respectively. These results suggested that Pib loci had different efficacy against blast infections in most rice-growing regions of Yunnan Province. Among 366 isolates, three genotypes (L1 with 1231bp, L2 with 3100bp, L3 with both of 1231bp and 3100bp) of AVR-Pib alleles in 162 isolates were amplified by AVR-Pib-specific primers (AVR-Pib F1/AVR-Pib R1) (Table 1; Additional file 4: Fig. S1), and the average percentage of the AVR-Pib allele was 44.3%. The highest percentage of amplification of AVR-Pib was 73.3% in the rice-blast isolates collected from northwestern Yunnan Province, whereas the lowest percentage was 36.1% from northeastern Yunnan Province (Table 1). The percentage of AVR-Pib was 46.3, 36.1, 73.3, 51.5, 67.9, and 39.0% in central, northeastern, northwestern, southeastern, southwestern and western Yunnan Province, respectively. The percentage of AVR-Pib was 47.0 and 42.4% in Xian/Indica (XI) rice- and Geng/Japonica (GJ) rice-production areas in Yunnan. The genotype of L1, L2 and L3 alleles of AVR-Pib was detected in 104, 53 and 5 isolates, with percentages of 28.4%, 14.5% and 1.3%, respectively (Table 1). The genotype of L1, L2 and L3 alleles of AVR-Pib was detected in the XI rice-production area, whereas L3 was absent in the GJ rice-production area (Table 1). These results suggested that the genome structure of AVR-Pib loci was complicated considerably in M. oryzae.
Table 1
Frequency of AVR-Pib genotypes and avirulent isolates of M. oryzae collected from Yunnan, China to IRBLb-B
|
Locations
|
No. of isolates
|
PCR detection
|
Pathogenicity assayb
|
|
Genotype and No. of isolates and frequency (%)
|
No. of avirulence isolates and Frequency (%)
|
|
L1
|
L2
|
L3
|
Total isolates and frequency (%)
|
|
Central
|
54
|
22 (40.7)
|
3 (5.6))
|
0
|
25 (46.3)B
|
41 (75.9)AB
|
|
Northeastern
|
72
|
23 (31.9)
|
3 (4.2)
|
0
|
26 (36.1)B
|
54 (75.0)B
|
|
Northwestern
|
15
|
11 (73.3)
|
0
|
0
|
11 (73.3)A
|
15 (100)A
|
|
Southeastern
|
33
|
10 (30.3)
|
6 (18.2)
|
1 (3.0)
|
17 (51.5)B
|
19 (57.6)C
|
|
Southwestern
|
28
|
9 (32.1)
|
8 (28.6)
|
2 (7.1)
|
19 (67.9A
|
15 (53.6)C
|
|
Western
|
164
|
29 (17.7)
|
33 (20.1)
|
2 (1.2)
|
64 (39.0)B
|
79 (48.2)C
|
|
Total
|
366
|
104 (28.4)
|
53 (14.5)
|
5 (1.4)
|
162 (44.3)
|
223 (60.9)
|
|
XI
|
149
|
31 (20.8)
|
34 (22.8)
|
5 (3.4)
|
70 (47.0)*
|
69 (46.3)**
|
|
GJ
|
217
|
73 (33.6)
|
19 (8.8)
|
0
|
92 (42.4)*
|
154 (71.0)**
|
|
Total
|
366
|
104 (28.4)
|
53 (14.5)
|
5 (1.4)
|
162 (44.3)
|
223 (60.9)
|
| a L1 indicates the AVR-Pib genotype with the expected size (1231bp), L2 and L3 indicates the AVR-Pib genotype with TE insertion (L2 with 3100bp, L3 with both of 1231bp and 3100bp). The superscript of A and B indicates the significant difference at 0.01 level, and * indicates non significant. |
| b Indicates pathogenicity assay on monogenic line IRBLb-B containing Pib. XI and GJ indicates Xian/Indica and Geng/Japonic, respectively. The superscript of A, B and C indicates the significant difference at 0.01 level, ** indicates significant difference at 0.01 level. |
Virulence function of AVR-Pib variations against the Pib gene
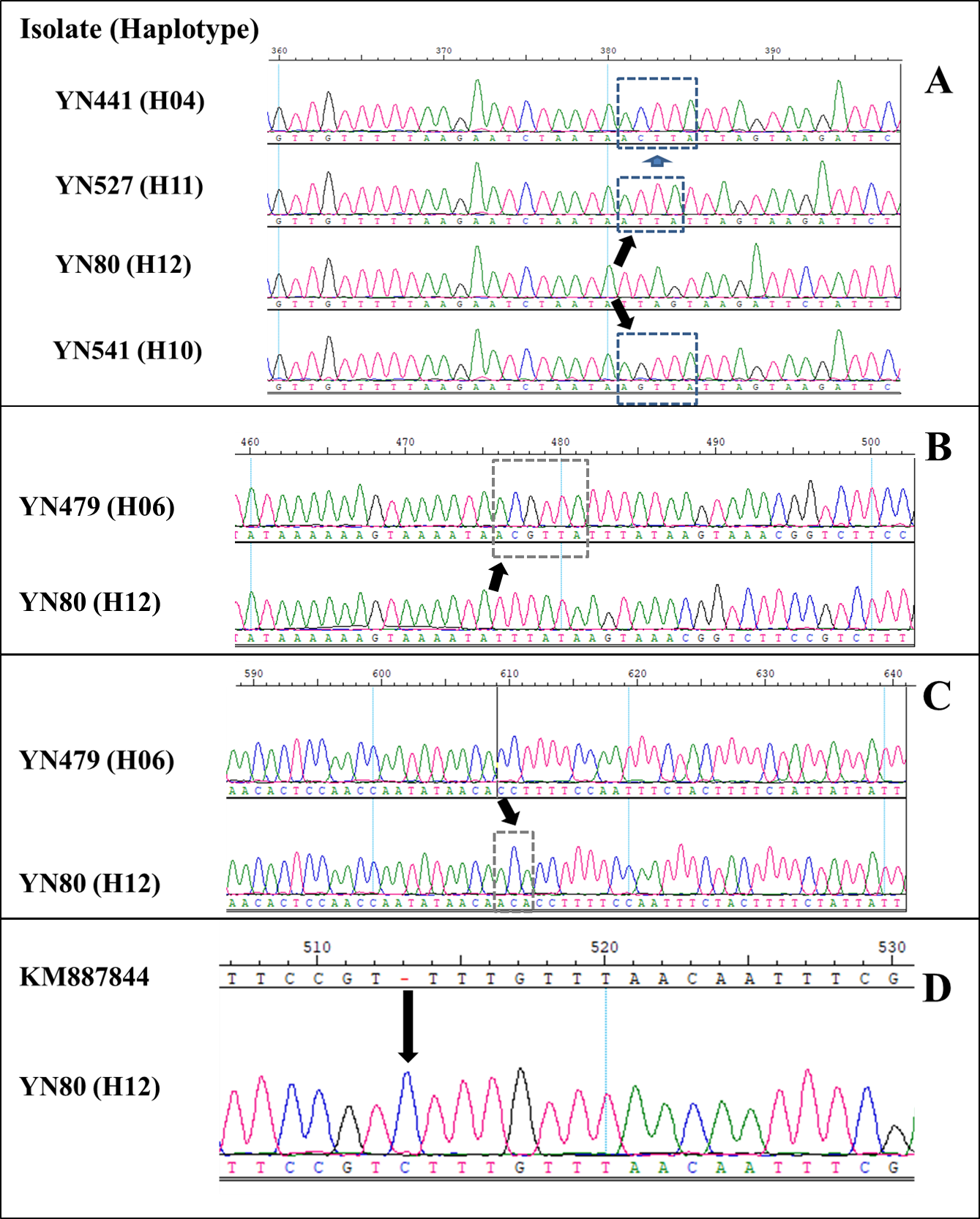
Twelve AVR-Pib haplotypes (H01 to H12) (Table 2), excluding the original AVR-Pib allele (GenBank accession number, KM887844), were detected on the nucleotide sequence assemblies of 90 isolates of L1 alleles containing a 719-bp 5′-region, 225-bp coding DNA sequence (CDS), and 302-bp 3′-region of AVR-Pib (Table 2; Additional file 5: Fig. S2). Also, insertion of Pot2 (at position -275) and Pot3 (at position -240) was identified based on the DNA sequence assemblies of six and 30 isolates (Table 2; Fig. 1), respectively, and the amplicon size difference between L1 and L2 (Additional file 4: Fig. S1). The 12 novel AVR-Pib haplotypes (H01-H12) were identified compared to previous published alleles [3, 27]. Alignment of DNA sequence assemblies of the AVR-Pib gene from 90 isolates revealed 18 mutation sites, including six mutant sites in the CDS region which weren’t in the signal-peptide region (Table 2; Additional file 5 and 6: Fig. S2 and S3). Six mutant sites in the CDS region led to changes in amino acids (Table 3). The CDS sequence assemblies of the AVR-Pib allele among the 126 isolates (including L1, L2 and L3) were predicted to produce four AVR-Pib proteins (Table 3). Among them, amino-acid variations were predicted to occur at six positions (Table 3). Amino-acid variations at F54L in H05 and H06, E46V, F53S, and F54V in H07 were found, these isolates of the corresponding haplotypes were avirulent on the monogenic line IRBLb-B (with Pib), whileas, the amino-acid variations at F47L, I49T, and R50G were found in one isolate with H08, which was virulent on the monogenic line IRBLb-B (with Pib) (Table 3), the amino-acid variations at F47, I49, and R50 in H01, H02, H03, H04, H05, H06, H07, H09, H10, and H12, and these isolates were avirulent on the monogenic line IRBLb-B (with Pib), whileas the amino-acid variations at 47L, 49T, and 50G in H08, and the isolate was virulent on IRBLB-b (Table 3). This finding suggested that the amino acids F47, I49, and R50 were crucial for the avirulence function of AVR-Pib.
Table 2
Haplotypes of AVR-Pib loci in M. oryzae field populations of Yunnan, China
|
Haplotype
|
No. of isolates
|
% of total
|
Variant locusa
|
|
5' UTR
|
CDS regions
|
3' UTR
|
|
-338
|
Between -325 and -326
|
Between -239 and -240
|
Between -216 and -217
|
-192
|
-175
|
Between -210 and -211
|
-93
|
137
|
141
|
146
|
148
|
158
|
160
|
+70
|
+154
|
+218
|
+232
|
|
KM887844
|
|
|
T
|
-
|
-
|
-
|
C
|
C
|
-
|
T
|
A
|
T
|
T
|
C
|
A
|
T
|
C
|
G
|
A
|
C
|
|
H01
|
33
|
26.2
|
.
|
.
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
H02
|
4
|
3.2
|
.
|
.
|
.
|
C
|
.
|
.
|
.
|
A
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
T
|
.
|
.
|
|
H03
|
4
|
3.2
|
.
|
.
|
.
|
C
|
T
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
H04
|
1
|
0.8
|
.
|
ACTTA
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
T
|
.
|
.
|
.
|
|
H05
|
8
|
6.3
|
C
|
.
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
C
|
.
|
.
|
.
|
.
|
|
H06
|
1
|
0.8
|
C
|
.
|
ACGTTA
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
C
|
.
|
.
|
.
|
.
|
|
H07
|
3
|
2.4
|
.
|
.
|
.
|
C
|
.
|
T
|
.
|
.
|
T
|
.
|
.
|
.
|
C
|
G
|
.
|
.
|
.
|
A
|
|
H08
|
1
|
0.8
|
.
|
.
|
.
|
C
|
.
|
.
|
ACA
|
.
|
.
|
A
|
C
|
G
|
.
|
.
|
.
|
.
|
.
|
.
|
|
H09
|
13
|
10.3
|
.
|
ACTTA
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
H10
|
10
|
7.9
|
.
|
AGTTA
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
H11
|
2
|
1.6
|
.
|
ATTA
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
H12
|
10
|
7.9
|
.
|
.
|
.
|
C
|
.
|
.
|
ACA
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
Pot2
|
6
|
4.8
|
-275 insert Pot2
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
Pot3 rev-A
|
1
|
0.8
|
-240 insert Pot3
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
Pot3 rev-B
|
2
|
1.6
|
-240 insert Pot3
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
C
|
.
|
.
|
.
|
.
|
|
Pot3-A
|
24
|
19.1
|
-240 insert Pot3
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
|
Pot3-B
|
2
|
1.6
|
-240 insert Pot3
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
C
|
.
|
.
|
.
|
.
|
|
Pot3-C
|
1
|
0.8
|
-240 insert Pot3
|
.
|
C
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
.
|
C
|
.
|
| a . Indicates the same with KM887844 (GenBank Accession No.). The KM887844 of AVR-Pib was obtained from GenBank. rev: indicates reverse insertion of Pot3 in AVR-Pib. |
Table 3
Variation of the AVR-Pib loci proteins in M. oryzae populations of Yunnan, China
|
Haplotype
|
No. of isolates
|
% of total
|
Variant locusa
|
Disease reactionb
|
|
46
|
47
|
49
|
50
|
53
|
54
|
|
KM887844
|
|
|
E
|
F
|
I
|
R
|
Y
|
F
|
|
H01
|
33
|
26.2
|
.
|
.
|
.
|
.
|
.
|
.
|
24R+5M+4?
|
|
H02
|
4
|
3.2
|
.
|
.
|
.
|
.
|
.
|
.
|
3R+1M
|
|
H03
|
4
|
3.2
|
.
|
.
|
.
|
.
|
.
|
.
|
4R
|
|
H04
|
1
|
0.8
|
.
|
.
|
.
|
.
|
.
|
.
|
1R
|
|
H05
|
8
|
6.3
|
.
|
.
|
.
|
.
|
.
|
L
|
7R+1?
|
|
H06
|
1
|
0.8
|
.
|
.
|
.
|
.
|
.
|
L
|
1R
|
|
H07
|
3
|
2.4
|
V
|
.
|
.
|
.
|
S
|
V
|
3R
|
|
H08
|
1
|
0.8
|
.
|
L
|
T
|
G
|
.
|
.
|
1S
|
|
H09
|
13
|
10.3
|
.
|
.
|
.
|
.
|
.
|
.
|
11R+2M
|
|
H10
|
10
|
7.9
|
.
|
.
|
.
|
.
|
.
|
.
|
9R+1M
|
|
H11
|
2
|
1.6
|
.
|
.
|
.
|
.
|
.
|
.
|
2S
|
|
H12
|
10
|
7.9
|
.
|
.
|
.
|
.
|
.
|
.
|
9R+1M
|
|
Pot2
|
6
|
4.8
|
.
|
.
|
.
|
.
|
.
|
.
|
7S
|
|
Pot3 rev-Ac
|
1
|
0.8
|
.
|
.
|
.
|
.
|
.
|
.
|
1S
|
|
Pot3 rev-B
|
2
|
1.6
|
.
|
.
|
.
|
.
|
.
|
L
|
2S
|
|
Pot3-A
|
24
|
19.1
|
.
|
.
|
.
|
.
|
.
|
.
|
22S+2M
|
|
Pot3-B
|
2
|
1.6
|
.
|
.
|
.
|
.
|
.
|
.
|
2S
|
|
Pot3-C
|
1
|
0.8
|
.
|
.
|
.
|
.
|
.
|
L
|
1S
|
| a. Indicates the same with KM887844. |
| b Indicates pathogenicity assay on the monogenic lines IRBLb-B containing the resistant gene of Pib. R, M and S indicate disease reaction was resistant, moderate resistant and susceptible, respectively; Ex. 24R indicated 24 isolates were avirulent to IRBLb-B; and ? indicates unknown. |
| c rev: indicates reverse insertion of Pot3 in AVR-Pib. |
The different haplotypes of H01, H02, H03, H04, H09, H10, H11 and H12 had no change on amino acids sequence (Table 3). The three-dimensional protein structures built by homology modeling (SWISS-MODEL; https://swissmodel.expasy.org/) showed the different protein structures of these four AVR-Pib variants (Additional file 7: Fig. S4). Isolates of H11 with insertion of ATTA in the 5′ UTR may change AVR-Pib expression and cause loss of the avirulent function (Tables 2 and 3; Additional file 6: Fig. S3). Isolates of H01 (amino acids same as that with a GenBank accession number of KM887844), H02, H03, H04, H05, H06, H07, H09, H10 and H12 haplotypes harbored AVR-Pib because these isolates were avirulent to the Pib-containing monogenic line IRBLb-B (Table 3). Isolate of H08 defeated the resistance of Pib because this isolate was virulent to the Pib-containing monogenic line IRBLb-B (Table 3). Furthermore, Pot2 and Pot3 inserted in the 5′ UTR of the Pib gene were identified in six isolates and 30 isolates (Table 3; Fig. 1), respectively, and these isolates were virulent to the Pib-containing monogenic line IRBLb-B (Table 3). These findings suggested that insertion of TEs (Pot2 and Pot3) and small segments of the nucleotide in the promoter region, and nuclear substitution in the ORF region, resulted in variation of AVR-Pib from avirulence to virulence, and that the diverse mutations of the AVR-Pib allele of M. oryzae were involved.
Haplotype diversity of AVR-Pib of M. oryzae
Among 12 AVR-Pib haplotypes, none were identical to the original AVR-Pib (GenBank accession number, KM887844) (Table 2). Eight haplotypes, as well as Pot2 and Pot3 insertion were detected in 50 M. oryzae isolates from western Yunnan Province (Table 4). Five haplotypes, as well as Pot3 and Pot3 reverse-insertion were identified in 23 M. oryzae isolates from central Yunnan Province. Three haplotypes, as well as Pot2 and Pot3 reverse-insertion were identified in 15 isolates of M. oryzae from northeastern Yunnan Province. Three haplotypes and Pot2 insertion were detected in 19 isolates from northeastern Yunnan Province. Three haplotypes and Pot3 insertion were identified in six M. oryzae isolates from southeastern Yunnan Province. Three haplotypes were identified in 13 isolates of M. oryzae from northwestern Yunnan Province (Table 4). Eleven and nine haplotypes were detected in GJ rice- and XI rice-production areas, and the diversity index (DI) of haplotypes was 0.84 and 0.79 for these areas, respectively. The DI of AVR-Pib was 0.72, 0.71, 0.70, 0.63, 0.59, and 0.54 for southeastern, central, western, southwestern, northeastern, and northwestern Yunnan Province, respectively (Table 4). In brief, the DI of AVR-Pib alleles in Yunnan Province was in the order southeastern > central > western > southwestern > northeastern > northwestern. The DI of AVR-Pib alleles in the GJ-rice production area was higher than that of in the XI rice-production area. These results indicate that the genetic divergence of AVR-Pib of M. oryzae in each rice growing region was occurred under fields condition.
Table 4
Haplotype distribution of AVR-Pib in different Yunnan rice growing regions
|
Haplotype
|
No. isolates
|
Percent (%)
|
Regions
|
Productionc
|
|
Central
|
Northeastern
|
Northwestern
|
Southwestern
|
Southeastern
|
Western
|
XI
|
GJ
|
|
H01
|
33
|
26.2
|
11(47.8)a
|
9(60.0)
|
2(15.4)
|
10(52.6)
|
0
|
1(2.0)
|
10(20.8)
|
23(29.5)
|
|
H02
|
4
|
3.2
|
0
|
0
|
0
|
4(21.1)
|
0
|
0
|
4(8.3)
|
0
|
|
H03
|
4
|
3.2
|
4(17.4)
|
0
|
0
|
0
|
0
|
0
|
0
|
4(5.1)
|
|
H04
|
1
|
0.8
|
0
|
0
|
0
|
0
|
0
|
1(2.0)
|
0
|
1(1.3)
|
|
H05
|
8
|
6.3
|
1(4.3)
|
1(6.7)
|
3(23.1)
|
0
|
2(33.3)
|
1(2.0)
|
2(4.2)
|
6(7.7)
|
|
H06
|
1
|
0.8
|
0
|
0
|
0
|
0
|
1(16.7)
|
0
|
1(2.1)
|
0
|
|
H07
|
3
|
2.4
|
0
|
0
|
0
|
0
|
0
|
3(6.0)
|
0
|
3(3.8)
|
|
H08
|
1
|
0.8
|
0
|
0
|
0
|
0
|
0
|
1(2.0)
|
1(2.1)
|
0
|
|
H09
|
13
|
10.3
|
2(8.7)
|
0
|
0
|
0
|
0
|
11(22.0)
|
0
|
13(16.7)
|
|
H10
|
10
|
7.9
|
2(8.7)
|
0
|
8(61.5)
|
0
|
0
|
0
|
0
|
10(12.8)
|
|
H11
|
2
|
1.6
|
0
|
0
|
0
|
0
|
1(16.7)
|
1(2.0)
|
1(2.1)
|
1(1.3)
|
|
H12
|
10
|
7.9
|
0
|
3(20.0)
|
0
|
1(5.3)
|
0
|
6(12.0)
|
6(12.5)
|
4(5.1)
|
|
Pot2
|
6
|
4.8
|
0
|
1(6.7)
|
0
|
4(21.1)
|
0
|
1(2.0)
|
6(12.5)
|
0
|
|
Pot3 rev
|
3
|
2.4
|
2(8.7)
|
1(6.7)
|
0
|
0
|
0
|
0
|
0
|
3(3.8)
|
|
Pot3
|
27
|
21.4
|
1(4.3)
|
0
|
0
|
0
|
2(33.3)
|
24(48.0)
|
17
|
10(12.8)
|
|
Total
|
126
|
100
|
23
|
15
|
13
|
19
|
6
|
50
|
48
|
78
|
|
No. of haplotypes
|
7
|
5
|
3
|
4
|
4
|
10
|
9
|
11
|
|
Index of diversityb
|
0.71
|
0.59
|
0.54
|
0.63
|
0.72
|
0.70
|
0.79
|
0.84
|
| a Number and frequency (in bracket) of isolates of each haplotype. |
| b Diversity index was calculated as the frequency of haplotypes types in the M. oryzae population following Fontaine’s method [43]. Diversity index = (1-∑ni=1pi2) (where pi is the frequency of the haplotype i in a population). |
| c XI and GJ indicates Xian/Indica and Geng/Japonic, respectively. |
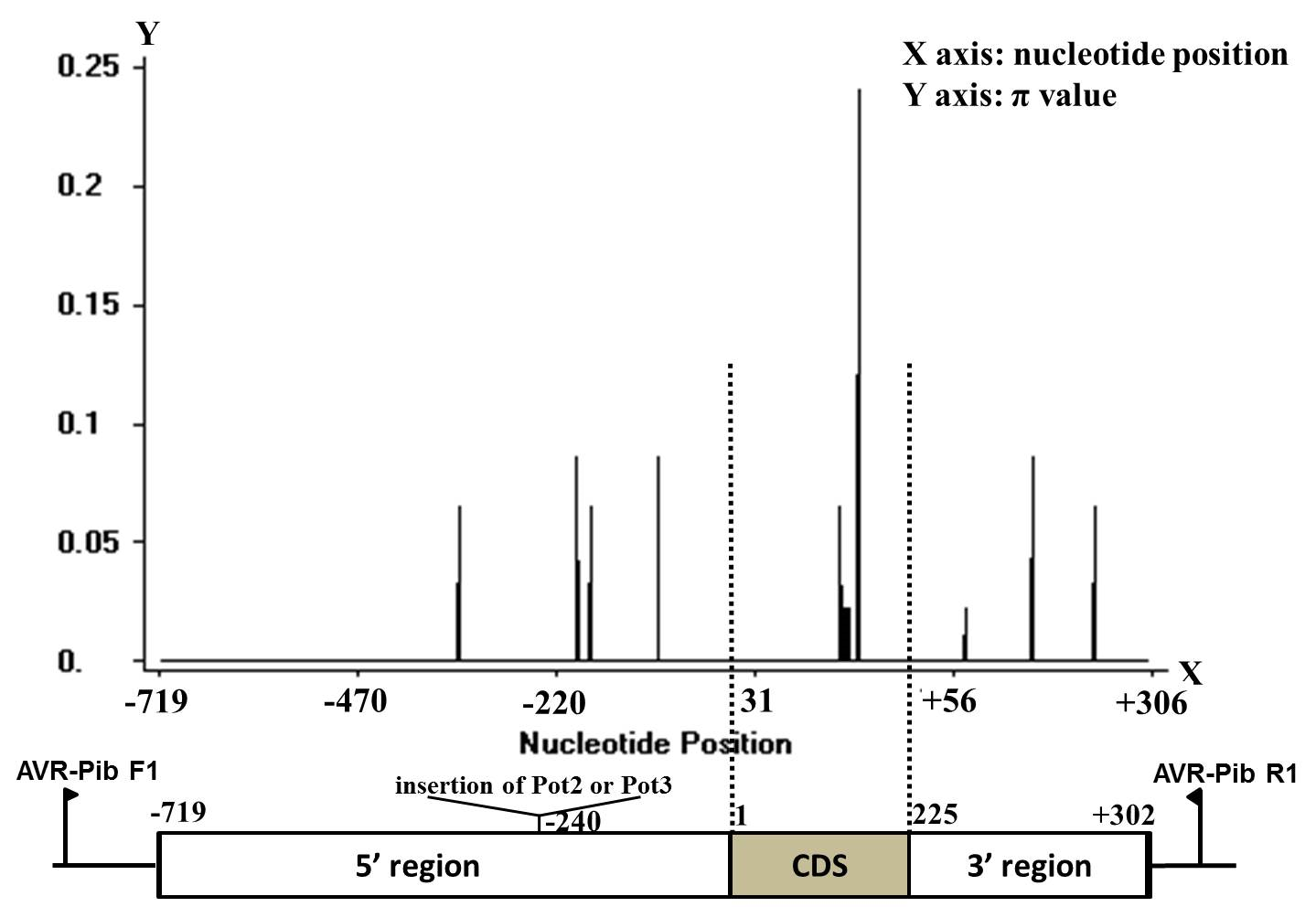
Eighteen nucleotide variable sites in AVR-Pib alleles were identified (Table 2; Additional file 5 and 6: Figs. S2 and S3). A haplotype network based on sequence variations of 90 isolates of L1 alleles was developed (Fig. 2). Four main lineage branches (A to D) of AVR-Pib were divided among 90 field isolates (Fig. 2), and different evolution of AVR-Pib among them was noted. Isolates of B and D lineage branches of AVR-Pib were avirulent to IRBLb-B (with Pib) (Fig. 2; Table 3). Isolates of H11 of the A-evolved branch and H08 of the C-evolved branch were virulent to the rice blast-resistant gene Pib, respectively (Fig. 2; Table 3). These data suggested that the A and C branches of AVR-Pib had evolved to virulence from avirulent origins via base substitution and insertion, and evaded the recognition of the rice blast-resistance gene Pib in field isolates. The virulence of H08 and H11 was identified in southeastern and western Yunnan Province (Table 4). Also, TE insertion in rice samples in all regions except northwestern Yunnan Province (Table 4) suggested that virulent evolution of AVR-Pib occurred in most rice-production areas of Yunnan Province.
Selection pressure on AVR-Pib in M. oryzae
The natural-selection pressure on AVR-Pib was calculated by Tajima's neutrality test on 126 AVR-Pib CDS sequences: the Tajima's D value was not significantly different from zero (D=−1.61687; 0.10>P>0.05) (Additional file 1: Table S1). This result suggested that AVR-Pib may suffer neutral selection and evolving neutrally in the population of M. oryzae. Furthermore, the results of three positive-selection models kept higher similatity (Additional file 8: Fig. S5). The “sliding window” under M8, M8a, and M7 models showed values of Ka/Ks (Ka, rate of nonsynonymous substitutions; Ks, rate of synonymous substitutions) across all 74 amino acids (Additional file 8: Fig. S5). The Ka/Ks value of all sites was >1 under M8 and M8a model, and the value was 1 under M7 model for entire residues, those results implying that these sites may have suffered neutral selection. These findings suggest that the AVR-Pib maybe under neutral evolution.
Adaption of TE insertion in AVR-Pib
We wished to confirm the host (rice and non rice) selection pressure on TE insertion in AVR-Pib. A total of 27 isolates from O. rufipogon (with Pib homologs) [20], Digitaria sanguinalis, Eleusine indica, E. coracana, and Musa nana Lour, which were stored in our lab, and 5 isolates genome sequence from Lolium perenne Linn (2 isolates), Setaria viridis (Linn.) Beauv. (1 isolate), and Triticum aestivum Linn (2 isolates) from Genbank were selected and analyzed (Additional file 2: Table S2). The AVR-Pib allele was not detected in the isolate from D. sanguinalis, M. nana Lour, Lolium perenne Linn, and Setaria viridis (Linn.) Beauv. (Additional file 2: Table S2). Only the L1 genotypes (with the expected size) of AVR-Pib were detected in isolates from E. indica, E. coracana, and Triticum aestivum Linn, suggesting that these isolates did not have a TE insertion in the AVR-Pib allele. Three genotypes of L1, L2 (with TE insertion), and L3 (with TE insertion) of AVR-Pib were detected in 18 isolates from the Pib homolog-containing O. rufipogon, and the isolate of YN441 (with the H9 haplotype of AVR-Pib and identical with the original haplotype of KM887844) was virulent to O. rufipogon (Fig. 3). These findings showed that the diversification of AVR-Pib of M. oryzae was dependent upon Pib homolog-containing O. rufipogon, and that variation in TE insertion in AVR-Pib could be selected and adapted to rice and other Gramineae species.
Phylogeny of Pib allele partial to CDS regions
Fifty-seven sequences of Pib were obtained from GenBank (Additional file 3: Table S3). Eleven of them were from five wild-rice species (seven from O. rufipogon, one from O. meyeriana, one from O. officinalis, one from O. longistaminata, one from O. nivara), and 46 accessions from O. sativa, including the original Pib (GenBank accession number, AB013448.1) (Additional file 3: Table S3). These sequences were aligned. A minimum-evolution phylogenetic tree was constructed based on the nucleotide sequences of exon 1 of Pib in 34 accessions and partial regions of exon-3 nucleotide sequences (from 7633 to 8484 of AB013448.1) of Pib in 49 accessions, respectively (Fig. 4). Exon 1 of Pib in wild-rice species (O. rufipogon, O. meyeriana, O. officinalis, O. longistaminata) was close to that in O. sativa (Fig. 4B). The DQ317978.1 group of the wild rice O. rufipogon shared >90% identity with the nucleotide sequences of the JN564624.1 group of Indica. Two major clades emerged in one part of exon 3 of Pib (Fig. 4C). One clade contained two wild-rice species (O. rufipogon and O. nivara) and O. sativa. The EF642422.1 group of the wild-rice species O. nivara shared >90% identity in nucleotide sequences with the EF642423.1 group of O. sativa. The EF642442.1 group of O. rufipogon shared >75% identity of nucleotide sequences with the EF642423.1 group of O. sativa. The other clade contained O. rufipogon and O. sativa. The EF642440.1 group of the wild-rice species O. rufipogon shared >75% identity of nucleotide sequences with the EF642433.1 group of O. sativa. The isolate of YN441 (with the H9 haplotype of AVR-Pib) was virulent to O. rufipogon, O. meyeriana, and O. officinalis (Fig. 3). These results suggested that different regions of the Pib gene may have suffered different selection pressures in the host rather than domestication.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}