Double lysogens of M. chelonae have increased resistance to antibiotics amikacin, kanamycin and tetracycline.
Wild type M. chelonae (CCUG 47445) carries a naturally occurring 67,657-bp prophage that we have named McProf. To determine how prophages impact gene expression and the antibiotic resistance phenotype of M. chelonae we added a second prophage. We identified three mycobacteriophages capable of infecting M. chelonae, Muddy, WildCat and BPs, of which only BPs is known to be temperate [22-24]. A double lysogen of M. chelonae was created from the WT M. chelonae strain using the cluster G mycobacteriophage, BPs [24]. BPs integrates into an attB site located within the 3' end of the host tRNA-Arg gene (BB28_RS01100), that is similar to the BPs attB site in M. smegmatis (Msmeg_6349) [24]. BPs lysogens of the M. chelonae WT strain (BPs, McProf) appear to be more stable than that of BPs lysogens of M. smegmatis. Lysogens form at a higher efficiency in M. chelonae WT (25%) compared to that in M. smegmatis (5%) and release fewer particles into cell culture supernatant (104 – 105 PFUs ml-1 compared to 1010 PFUs ml-1) [24, 25].
To determine if the presence of a second prophage in M. chelonae alters susceptibility to antibiotics, we tested the viability of the double M. chelonae lysogen (BPs, McProf) relative to the WT strain (M. chelonae +McProf) in the presence of varying levels of the aminoglycosides, amikacin (AMK) and kanamycin (KAN), and tetracycline (TET) (Fig. 1). The presence of the second prophage, BPs, significantly increased resistance to both aminoglycosides and the tetracycline. As a positive control, we exposed the M. chelonae (McProf) strain to sub-inhibitory concentrations of acivicin (ACI), a known inducer of intrinsic resistance in mycobacteria [10]. As expected, ACI significantly increased the viability of M. chelonae (McProf) in both the AMK and KAN assays (Fig. 1a and 1b). The effect of ACI on M. chelonae (McProf) viability in the TET assay was less dramatic, with only a slight increase in viability relative to M. chelonae (McProf) in the absence of ACI. The presence of the second prophage, BPs, had a greater effect on TET resistance than the ACI.
Isolation of a non-lysogen and single BPs lysogen of M. chelonae.
To better understand how the presence of the second prophage increases antibiotic resistance, we generated a strain of M. chelonae that contains no prophage (M. chelonae (ΔMcProf)) and from that a single BPs lysogen of M. chelonae. To remove the McProf prophage we created a recombinant strain of M. chelonae (McProf) that overexpresses the McProf excise gene, gp5, from an inducible mycobacterial expression plasmid (Table 1) [26]. Using sets of PCR primers that amplify either the bacterial attachment site (attB) and the phage attachment site (attP) or the hybrid prophage attachment sites, attL and attR, we identified ATc-induced bacterial colonies that had an intact attB site, indicating that the McProf prophage had been lost and that McProf has an active integrase system (Table 2) (data not shown). To determine if McProf phage particles are released from M. chelonae (McProf) cells through spontaneous induction, concentrated culture supernatants were plated onto lawns of the newly acquired non-lysogen strain (ΔMcProf), but we were unable to detect plaques. PCR analysis of M. chelonae (McProf) culture supernatants also failed to detect the McProf attP sequence, which would have indicated the presence of either excised McProf genome or linear McProf genome in phage particles. It is possible that there is a mutation that we were not able to identify that prevents McProf from carrying out a successful lytic infection. Alternatively, M. chelonae may not be the natural host and McProf is capable of lytically infecting other mycobacterial hosts.
The non-lysogen strain of M. chelonae (ΔMcProf) was used to isolate single lysogens of BPs. Although we were able to isolate BPs lysogens in the non-lysogen strain of M. chelonae, they are less stable than BPs lysogens formed in the WT strain (McProf) and comparable to lysogens formed in M. smegmatis [24, 25]. Lysogens formed at an efficiency of ~5%, and the titer of BPs in lysogen culture supernatants was 1010 PFUs ml-1, several orders of magnitude higher than that of the double lysogen (105 PFUs ml-1).
Single and double lysogens carrying McProf have higher AMK resistance than strains that lack McProf.
To determine the roles of prophages BPs and McProf in the increased resistance observed in the double lysogen, we tested the viability of double (BPs, McProf) and single (BPs or McProf) M. chelonae lysogens relative to non-lysogen cells (ΔMcProf) in the presence of varying levels of AMK and TET (Fig. 2). Because AMK and clarithromycin (CLA) are the major drugs used to treat M. abscessus infections, we focused on AMK over KAN and added an assay with the macrolide, CLA. The presence of the naturally occurring prophage, McProf, significantly contributes to AMK resistance in M. chelonae (Fig. 2a). The WT strain carrying McProf alone had higher growth at 30 µg mL-1 than the non-lysogen strain (ΔMcProf) that had been treated with sub-inhibitory concentrations of ACI, a known inducer of whiB7 and intrinsic aminoglycoside resistance [8, 10].The presence of a second prophage, BPs, further increases resistance to AMK, with bacterial growth at doses as high as 60 µg mL-1 and growth approached that of WT strains (McProf) treated with ACI. BPs alone had no effect on AMK resistance suggesting that BPs only increases AMK resistance through an interaction with the naturally occurring prophage, McProf.
A slight increase in TET resistance in M. chelonae required the presence of both prophages (Fig. 2b). Double lysogens (BPs, McProf) were more resistant to TET at doses of 16 and 8 µg mL-1, than single lysogens (McProf or BPs) and non-lysogens (ΔΔMcProf) whereas the presence of McProf alone had no effect on TET. ACI also did not induce significant TET resistance in WT (McProf) and non-lysogen (ΔMcProf) strains.
The presence of prophages had a subtler effect on CLA resistance, which was not surprising as M. chelonae lacks the gene erm, which provides macrolide resistance in M. tuberculosis and M. abscessus (Fig. 2c.) [27]. Only the double lysogen demonstrated some difference in CLA resistance relative to M. chelonae (McProf) but at a very low dose of 0.09 µg mL-1.
Prophage McProf enhances AMK resistance in response to sub-inhibitory concentrations of antibiotics.
Because the M. chelonae (McProf) strain treated with ACI had higher AMK resistance than the non-lysogen strain treated with ACI, we wondered if the presence of prophage McProf enhances the effect of sub-inhibitory concentrations of antibiotics on AMK resistance. To determine the interaction between ACI and the presence of one or both prophages, we pre-treated each of the four lysogen and non-lysogen strains with sub-inhibitory concentrations of ACI and repeated the AMK resistance assay. The presence of McProf increases the effect of ACI on AMK resistance compared to the non-lysogen whereas the BPs prophage alone does not (Fig. 3). The double lysogen treated with ACI had the highest AMK resistance suggesting that both BPs and ACI interact with McProf to increase AMK resistance.
The whiB7 regulon is upregulated in double lysogens of M. chelonae.
RNAseq analysis was performed on RNA isolated from the WT (McProf) and double lysogen (BPs, McProf) M. chelonae strains to learn if the presence of the second prophage, BPs, impacted expression of genes that may be involved in mycobacterial antibiotic resistance. The presence of prophage BPs significantly altered expression of M. chelonae genes, including numerous putative virulence genes. Out of 4,867 genes in the M. chelonae genome, 417 (8.5%) were differentially regulated in the double lysogen (BPs, McProf) (Fig. 4).
The majority of the top-ranked genes in the double lysogen belonged to the whiB7 regulon, genes in M. tuberculosis with functions related to antibiotic resistance and increased survival in macrophage (Tables 3 and 4) [28]. The transcription factor, identified as whiB7 (BB28_RS17590), was the fifth most highly upregulated gene in the double lysogen with a fold change of 26.5 (Log2FC = 4.7, FDR = 1.3–73) (Table 3). The WhiB7 peptide sequence shares 95% identity with the M. abscessus WhiB7 peptide (MAB_3508c) and has all the conserved residues that form the iron sulfur cluster binding domain. It also has the glycine-rich motif of the signature WhiB7 C-terminal “A/T Hook” DNA binding domain, which binds to AT-rich sequences adjacent to target gene promoters [9, 10]. The M. chelonae genome contains a large whiB7 regulon like that of M. abscessus, with 103 of the 128 whiB7 regulon genes found in M. abscessus [10]. We identified a total of 30 upregulated genes that belong to the whiB7 regulon, which included GNAT acetyltransferases, eis1 (BB28_RS05390) and eis2 (BB28_RS22650), multi-drug efflux transporter tap (BB28_RS06750), the tetV efflux pump (BB28_RS13560) and numerous ABC transporters with ATP binding domains that possibly function as efflux pumps (Tables 3 and 4). In M. abscessus and M. tuberculosis, erm provides macrolide resistance but the gene is not present in the M. chelonae genome [27]. Although M. chelonae lacks this whiB7 regulon gene, there was still a slight increase in CLA resistance in the double lysogen relative to the M. chelonae (McProf) strain [27]. This small difference in resistance may be due to expression of a newly discovered gene in the whiB7 regulon, the ribosome splitting factor hflX (MAB_3042c), which is reported to contribute to macrolide resistance in M. abscessus [29]. Expression of the M. chelonaehflX was slightly elevated in double lysogens relative to the WT strain (McProf) (BB28_RS14985; Log2FC=1.5, FDR=8.4-33) (Table 4). An additional 25 whiB7 regulon genes were upregulated but had fold changes of less than 2.
The most highly regulated gene, with a 99-fold increase in expression, was annotated as a flotillin protein with no known function (BB28_RS01845) (log2FC = 6.6, FDR = 6.4–124) (Table 3). Several of the most down regulated genes in the lysogen include a padR-family transcription factor (BB28_RS01835, Log2FC= -8.3, FDR = 1.4–10) and genes involved in glycerol uptake (glpF, BB28_RS01835, Log2FC= -6.2, FDR = 4.5–06) and metabolism (glpK, BB28_RS01840, Log2FC= -9.0, FDR = 1.1–12)
(Table 3) [30].
Upregulation of whiB7 only occurs in double lysogens of M. chelonae.
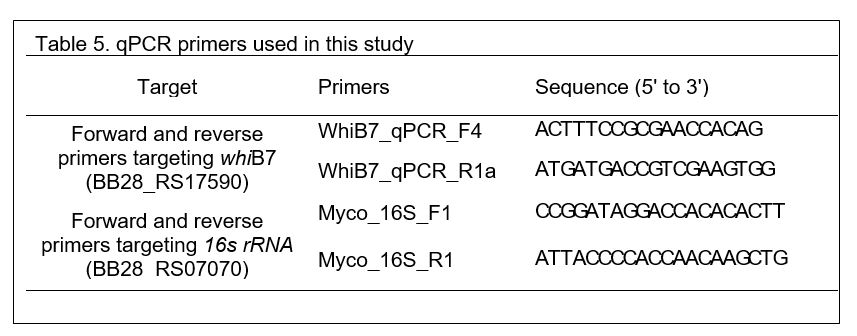
To determine how the presence and absence of each prophage impacts whiB7 expression, whiB7 mRNA levels were measured by qPCR in the BPs single lysogen, double lysogen (BPs, McProf), and non-lysogen (ΔMcProf) and compared to that of the WT strain (McProf) (Fig. 5). Although whiB7 expression was slightly elevated in the non-lysogen (2-fold) and BPs single lysogen (4-fold) strains relative to the WT strain (McProf), the dramatic increase in whiB7 expression (~40-fold) only occurred in M. chelonae carrying both prophages (BPs, McProf). The elevated whiB7 expression occurred in the absence of known inducers of whiB7, such as ACI, which suggests BPs interacts with prophage McProf, resulting in whiB7 induction. The elevated expression of whiB7 in the double lysogen likely explains the large increased resistance to AMK and the smaller increases in resistance to TET and CLA in the absence of ACI treatment (Fig. 2).
Sub-lethal concentrations of ACI but not AMK induce whiB7 expression in the double lysogen of M. chelonae.
We were surprised that the M. chelonae (McProf) strain had the lowest expression of whiB7 expression among the four strains given that it demonstrated the second highest AMK resistance, both in the presence and absence of ACI. We reasoned this may be due to whiB7-independent intrinsic resistance, such as cell wall permeability, and/or whiB7 induction in the presence of AMK, which is a more potent inducer of whiB7 than ACI [10]. Likewise, we wondered if the heightened AMK resistance observed in the single and double McProf lysogen strains in the presence of ACI was due to increased whiB7 expression. We therefore measured whiB7 expression in all four strains in the presence and absence of sublethal concentrations of ACI (75 μM) or AMK (16.7 μM) (Fig. 6).
ACI treatment resulted in increased whiB7 expression in all four strains relative to untreated strains (Fig. 6a). Expression of whiB7 was highest in the double lysogen strain (BPs, McProf) treated with ACI which correlates with the observed AMK resistance of this strain. whiB7 expression in the single and non-lysogen strain increased with ACI treatment; however, the relative levels of whiB7 expression did not correlate with AMK resistance (Fig. 2a). Although the fold-increase of whiB7 in ACI-treated strains relative to control strains was highest in the WT strain (McProf) (9.5-fold) whiB7 was lower than that of the ACI-treated BPs single lysogen, which demonstrated lower AMK resistance.
To determine if exposure to AMK also contributes to whiB7 expression in each of the four strains, whiB7 expression was determined in each of the strains in the presence and absence of sub-lethal concentrations of AMK (16.7 μM). Strains that lack the McProf prophage had the greatest increase in whiB7 expression in response to AMK treatment. The non-lysogen and BPs single lysogen had 28- and 7-fold increases in whiB7 expression in response to AMK treatment, respectively (Fig. 6b). AMK had less of an effect on whiB7 expression in strains carrying McProf. AMK treatment resulted in a 3.5-fold increase in whiB7 expression in the single McProf lysogen and no significant increase in whiB7 expression in the double lysogen. It’s possible that AMK doesn’t result in strong induction of whiB7 expression in the McProf+ strains due to cell wall permeability and/or efflux, and if so, this could also explain the AMK-resistant phenotypes observed in the single McProf and double lysogen strains.
Organization of the McProf prophage genome.
To better understand how the two prophages, BPs, and McProf, may be interacting to alter whiB7 expression, we characterized the McProf genome and examined viral gene expression profiles from both McProf and BPs prophage genomes in the double lysogen. The McProf genome is 67,657 bp in length (M. chelonae CCUG 47445 coordinates 1,521,426 – 1,589,648) and encodes 98 putative genes and no tRNAs (Fig. 7a). The prophage genome is flanked by 45-bp phage attachment sites, attL and attR (5'- TGCGCCGTCAGGGGCTCGAACCCCGGACCCGCTGATTAAGAGTCA). The right attachment site, attR, overlaps a leftward oriented tRNA-Lys (BB28_RS07905). Located adjacent to the left attachment site, attL, is a rightward transcribed tyrosine integrase (gp1), one gene of unknown function (gp2) and a leftward transcribed gene, gp3, that is likely to be the immunity repressor, as it shares high amino acid sequence similarity with the immunity repressors of singleton mycobacteriophage DS6A (66%) and cluster K2 mycobacteriophages (70%) DismalFunk, DismalStressor, Findley, Marcoliusprime and Milly [31]. Gp4 and gp5 both have helix-turn-helix DNA binding motifs and encode Cro (control of repressor’s operator) and excise, respectively.
Located between attR and the structural genes (gp51 – 82) are genes that are typically expressed during lysogeny [32]. We were unable to predict a function for the majority of these genes; however, we were able to identify an ADP-riboysl glycosylhydrolase (gp86), a helix-turn-helix DNA binding protein (gp89), a membrane protein (gp90), and an AAA-ATPase (gp91). Most intriguing is the leftward transcribed gene cassette immediately adjacent to attR, which encodes proteins that may be secreted by the mycobacterial Type 7 secretion system (T7SS) (Esx-3 or Esx4) (Fig. 7b and c). Gp98 encodes a 105-amino acid gene product that forms four HHpred predicted helical domains with high probability matches to WXG-100 family motifs of T7SS proteins. The gp98 sequence contains a SAG motif, which strays slightly from the conserved WXG motif that is characteristic of T7SS secreted substrates [33]. Gp97 encodes a 732-residue polymorphic toxin that has a WXG-100 motif in the N-terminus and a possible T7SS secretion signal (YxxxD/E) in the C-terminus [34]. The C-terminus also includes a toxin_43 motif (PF15604.6) and high sequence similarity to the C-terminus of Type 6 secretion system (T6SS) polymorphic toxin, TdeI (Atu4350), found in Agrobacterium tumefaciens [35]. This family of proteins has DNAse activity and shares a conserved HXXD catalytic domain located in the C-terminus (Fig. 7b) [35]. Tde toxins are typically paired with a Tdi immunity protein and a likely immunity protein, gp96, was identified downstream of McProf gp97. McProf gp96 encodes a putative 216-residue protein that contains GAD-like and DUF1851 domains, which are well-conserved domains of Tdi homologs (Fig. 7c) [35].
Although the McProf prophage was identified and characterized in M. chelonae, it is closely related to prophages found in clinical M. abscessus isolates. BlastN analysis of the McProf prophage genome in M. abscessus-specific databases (e.g. phagesdb.org) identified 25 M. abscessus isolates with McProf-like prophage sequences [31]. The WXG-100 family polymorphic toxin cassette identified in McProf is also prevalent in M. abscessus genomes. BlastP analysis of the McProf Tde-like polymorphic toxin (gp97 toxin) results in 100 high-similarity protein alignments to mycobacterial proteins with 91% matching M. abscessus sequences. An initial random screen of 10 of the aligned sequences showed that they were all located in prophage genomes flanked by a WXG-100 family gene and an immunity gene.
Lysogenic gene expression profiles from the BPs and McProf prophage genomes.
To determine if the presence of BPs alters gene expression from the McProf prophage genome, differential expression of McProf genes was examined between the WT strain (McProf) and the double lysogen (BPs, McProf). None of the expressed McProf genes were significantly differentially expressed in the presence of the BPs prophage (FC>1.99 and FDR<0.05). Because there was no difference in expression profiles, we present below the expression profile of only the McProf prophage from the M. chelonae WT (McProf) strain (Fig. 8b).
The immunity repressors from both the BPs (gp33) and McProf (gp3) genomes are highly expressed during lysogeny of M. chelonae (Fig. 8a). The BPs genome also expresses, gp58, a gene of unknown function that is part of a mycobacteriophage mobile element (MPME1) (Fig. 8a) [24]. There are an additional 15 genes expressed at varying levels from the McProf genome (Fig. 8b). The integrase (gp1) is expressed at low levels and is adjacent to a moderately expressed genes of no known function (gp2). There are three reverse oriented genes, gp48 – 50, located between the HNH endonuclease (gp47) and the small subunit terminase (gp51) and a small reverse oriented gene (gp56) adjacent to the scaffolding protein with moderately and low expression, respectively. We were not able to determine functions for these genes; however, gp46 and gp47 do have predicted membrane domains. The remaining genes expressed from the McProf prophage genome are located between the structural genes and attR and many do not have predicted gene functions, including the most highly expressed McProf gene, gp84. There is also strong expression from the gene cassette containing the putative WXG-100 family polymorphic toxin and immunity protein (gp96 – 98). None of the expressed McProf genes were significantly differentially expressed in the presence of the BPs prophage (FC>1.99 and FDR<0.05).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}