Titanium dioxide nanoparticles (TiO2 NPs) have been widely used in various industries, such as chemistry, paint, food, and medicine [23]. It has been reported that TiO2 NPs induce central nervous system (CNS) dysfunction by accumulating in the brain [24]. In zebrafish, a low dose of TiO2 NPs (5 µg/L) can lead to the aggregation of TiO2 NPs in neurons, the increase of apoptosis, and the decrease of spatial recognition [25]. In vitro, TiO2 NPs were also found to induce apoptosis in primary cultured hippocampal neurons [12]. In the process of TiO2 NPs-induced neuron apoptosis, it was found that TiO2 NPs could promote the decrease of ΔΨm, the release of cytochrome c, and the increase of caspase-3 level [24]. Those results indicated that mitochondria participate in the apoptosis of neurons induced by TiO2 NPs. Mitochondria are semi-autonomous organelles responsible for energy production, cell differentiation, cell information transmission, cell growth, and cell cycle [26]. Given their many crucial functions in cell physiology, it is not surprising that mitochondria are involved in, or mediate, cognitive deficits, even before neuronal damage. Although some studies have reported the damaging effect of TiO2 NPs on mitochondrial function, to date, most studies have focused on the downstream signaling pathway after mitochondrial function damage, such as the activation of the apoptosis pathway or oxidative stress effects. The molecular mechanisms of mitochondrial function regulation under the influence of TiO2 NPs remain unclear. Understanding these mechanisms is essential to protect CNS function under TiO2 NPs exposure. In the present study, we demonstrated that TiO2 NPs are strong positive regulators of miR-21-5p expression, which protects the neuron cells in response to TiO2 NPs, and that VDAC1 plays an important role in the function of miR-21-5p by preserving oxygen consumption rate, ΔΨm, and the ATP content.
MiR-21 is considered an oncogenic molecular due to inhibiting the expression of phosphatases, which regulate the signaling pathways such as AKT and MAPK in cells [27, 28]. In non-cancer cells, miR-21 usually acts as a protector [29, 30, 31]. Although microRNA-21 (miR-21) is believed to be related to the regulation of mitochondrial energy metabolism [32, 33, 34, 35], the effect of TiO2 NPs on miR-21 has not been reported. In the present study, we discovered that the level of miR-21-5p was upregulated in primary neuron cells, SH-SY5Y cells, and rat prefrontal cortex under TiO2 NPs treatment in a dose- and time-dependent manner (Fig. 1A − 1F). Considering that TiO2 NPs also led to a decrease in the ΔΨm (Fig. 1G) and ATP content (Fig. 1H), miR-21-5p seemed to be a cause or result of mitochondrial dysfunction under TiO2 NPs treatment. To confirm the function of miR-21-5p, the correlation of the miR-21-5p level with the ΔΨm or ATP content of SH-SY5Y cells treated with TiO2 NPs was analyzed, and SH-SY5Y cells (miR-21) and SH-SY5Y cells (shmiR-21) were employed (Fig. 2C &2D). Under TiO2 NPs treatment, there was a significant correlation between miR-21-5p and the ΔΨm (Fig. 2A) or the ATP content (Fig. 2B). Knockdown of miR-21-5p resulted in a significant decrease in the ΔΨm and ATP content of SH-SY5Y cells under TiO2 NPs treatment. Overexpression of miR-21-5p resulted a significant increase in the ΔΨm and ATP content of SH-SY5Y cells under TiO2 NPs treatment (Fig. 2E & 2F). Our results indicated that the increase of miR-21-5p expression induced by TiO2 NPs is an important protective mechanism to preserve the ΔΨm and ATP content in neuron cells.
In the process of mitochondrial electron transport, the electrochemical potential energy is stored in the inner membrane of mitochondria. On both sides of the inner membrane, the asymmetric distribution of proton and other ion concentration will form the ΔΨm, which is the core driving oxidative phosphorylation to produce ATP [36]. The oxygen consumption rate (OCR) is the basis of mitochondrial oxidative phosphorylation and ATP synthesis [14, 37]. In our study, OCR was measured using the Seahorse system in SH-SY5Y cells (shmiR-21) and SH-SY5Y cells (Nonsense) under TiO2 NPs treatment. Basal, ATP-linked, and maximal respiration were remarkably declined in SH-SY5Y cells (shmiR-21) compared with those in SH-SY5Y cells (Nonsense) (Fig. 3B). This inhibition in ATP-linked respiration corresponded with the decreased ATP content in SH-SY5Y cells (shmiR-21) (Fig. 2F). Thus the damaged OCR induced by TiO2 NPs was protected by mir-21-5p. Although glycogen depletion, caused by uncontrolled glycolysis, leads to cognitive impairment [38], glycolysis is the main source of energy supply for anoxic neurons. Moreover, previous studies suggested that lactate, a glycolytic product, plays an important role in memory processing [14]. Our results showed that miR-21 knockdown reduced the extracellular acidification rate (ECAR) induced by oligomycin under TiO2 NPs treatment (Fig. 3C). This suggested that glycolysis is also one of the ways by which miR-21 protects neurons under TiO2 NPs treatment.
Voltage-dependent ion channels are involved in the process of energy metabolism [39]. Here, we determined the expression levels of VDACs, and found that VDAC1 expression was regulated by miR-21 (Fig. 4A) as well as by TiO2 NPs (Fig. 4B). VDAC1plays an important role in glycolysis and oxidative phosphorylation [40]. High levels of VDAC1 were demonstrated in the brains of patients with Alzheimer’s disease and in amyloid precursor protein transgenic mice [41]. Some studies have found that Vdac1 overexpression is related to the release of cytochrome c [40]. However, in the present study, VDAC1 overexpression reversed the reduction of the ΔΨm (Fig. 4D) and ATP content (Fig. 4E) induced by miR-21 knockdown in SH-SY5Y cells under TiO2 NPs treatment. Basal, ATP-linked, and maximal respiration also increased significantly in SH-SY5Y cells (VDAC1, shmiR-21) compared with those in control SH-SY5Y cells (Vector, shmiR-21) (Fig. 4F). Therefore, we speculated that TiO2 NPs would upregulate the VDAC1 via miR-21 accumulation, which, as an endogenous protection mechanism, consequently preserves the OCR, ΔΨm, and ATP content.
Up to now, no mechanism of titanium dioxide regulating VDAC has been reported. To explore how miR-21 regulates VDAC1 under TiO2 NPs treatment, we detected the VDAC1 mRNA level in SH-SY5Y, SH-SY5Y (miR-21), and SH-SY5Y (shmiR-21) cells with TiO2 NPs treatment (Fig. 5A − 5C). Thus, miR-21 might upregulate the VDAC1 by promoting its mRNA transcription under TiO2 NPs treatment. To verify this speculation, the promoter-dependent transcriptional activities of VDAC1 were detected in SH-SY5Y cells with or without TiO2 NPs treatment. The results indicated that VDAC1 promoter transcription factor binding sites (-908 to -2000) were responsible for VDAC1 mRNA upregulation in TiO2 NPs-treated cells (Fig. 5D & 5E). Although studies have reported that the transcriptional regulation of VDAC1 plays an important role in different cell models [42, 43, 44], the mechanism of VDAC1 transcription regulation is still unclear, especially under the intervention of TiO2 NPs. Bioinformatic methods predicted transcription factor binding sites and then the levels of the predicted transcription factors using western blotting in cells with or without TiO2 NPs treatment (Fig. 5G). The level of phosphorylated (P)-c-Jun (Ser73) was upregulated markedly in TiO2 NPstreated SH-SY5Y cells compared with that in the control cells. Furthermore, inhibition of c-Jun phosphorylation reversed the upregulation of VDAC1 induced by TiO2 NPs and decreased the ΔΨm and ATP content in SH-SY5Y cells. Based on these results, we proposed a previously unreported mechanism of VDAC1 transcriptional regulation and the protective role of VDAC1 protein in neuron cells in response to TiO2 NPs.
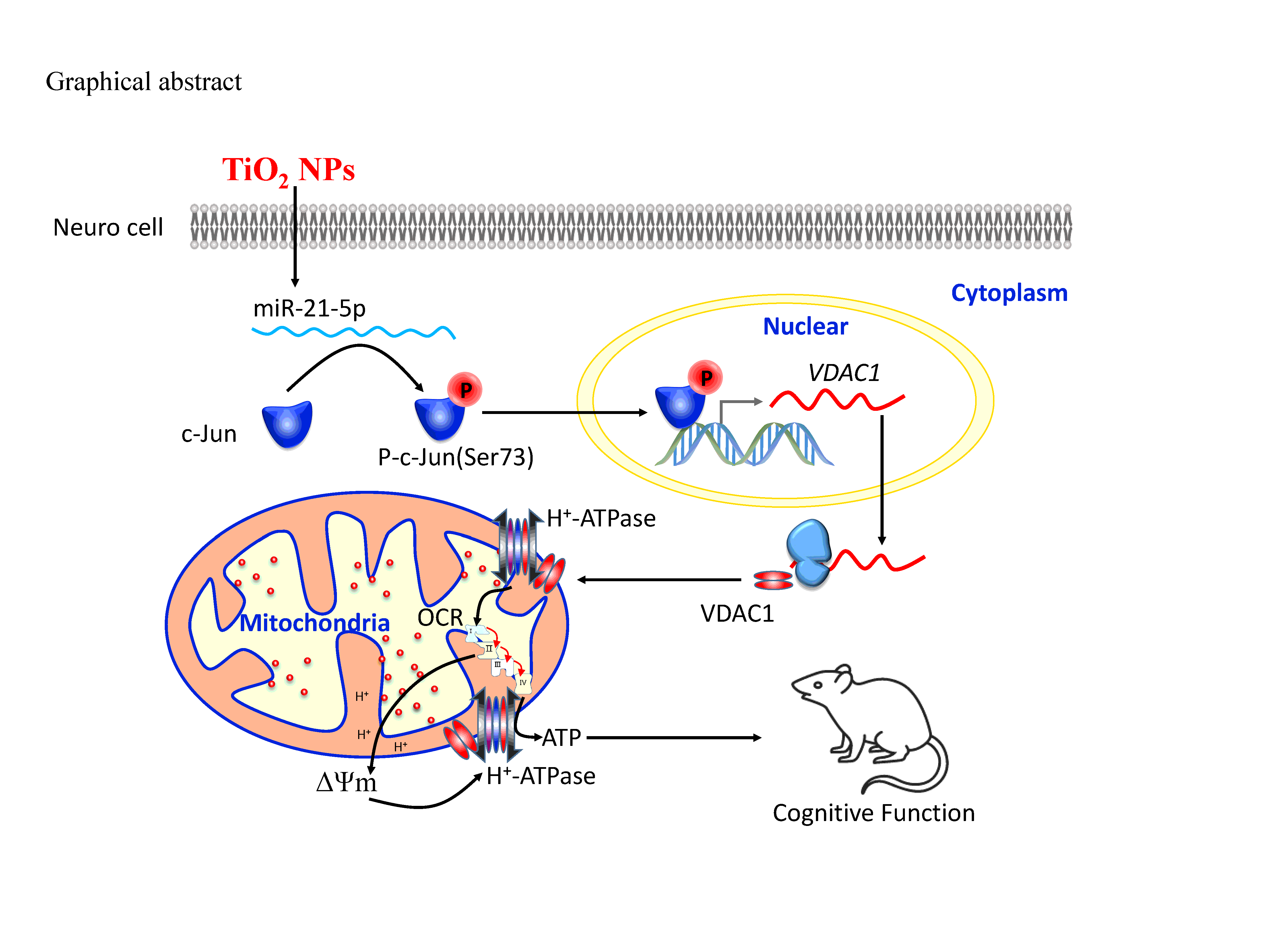
In summary (Graphical abstract), our results defined a novel effect of TiO2 NPs on miR-21-5p upregulation, which would inhibit the expression of an as yet unidentified phosphatase, leading to increased or maintained phosphorylation of c-Jun. This active form of c-Jun would increase the transcriptional activity of VDAC1 leading to the upregulation of VDAC1 protein levels, an increased OCR, elevation of the ΔΨm, and preservation of the ATP content. These findings demonstrated the protective of miR-21 in TiO2 NPs-induced neuron cells, providing novel insights into the multiple levels of VDAC1 protein regulation and revealed an endogenous neuroprotective mechanism in response to TiO2 NPs.
{kind=link}