RNF8 is a 484-amino-acid E3 ligase located on chromosome 6p21.3(1). as demonstrated previously, the biological function of RNF8 largely originates from its two conserved domains: the FHA domain in the N-terminal, which binds to the phospho-peptides motif, and the RING domain in the C-terminal to catalyze the formation of the poly-ubiquitin chain(16). As an E3 ligase, RNF8 has been shown to promote the formation of K63, K48, and K11-linked polyubiquitin chains when coupled with E2s such as UBC13, (UBCH8, UbcH6, UBE2E3), and UBE2S, respectively. These ubiquitination on RNF8-interacting targets would contribute to their nuclear translocation, activation, protein degradation, etc(6, 40). Based on these functions, RNF8 were played important roles in various biological processes like DNA damage response, telomere protection, cell cycle control, and transcriptional regulation(5). However, it is quite confusing that RNF8 seems to have two faces. One is that RNF8 acts as a “Guardian” of our cell, it helps transduce DNA damage signals and initiate DSBs repair upon DNA damage, maintain genomic stability and participate in spermatogenesis(12). Down-modulation of RNF8 also enhances cancer cell radiosensitivity(41). While another face of RNF8 is to promote lung cancer tumorigenesis, chemoresistance(2, 35), and promote breast cancer metastasis(3), like a “Villain”. In light of the structure of RNF8 and the way RNF8 functions, we utilize LC-MS to identify its direct targets and interacting proteins in order to understand how RNF8 participates in distinct biological processes.
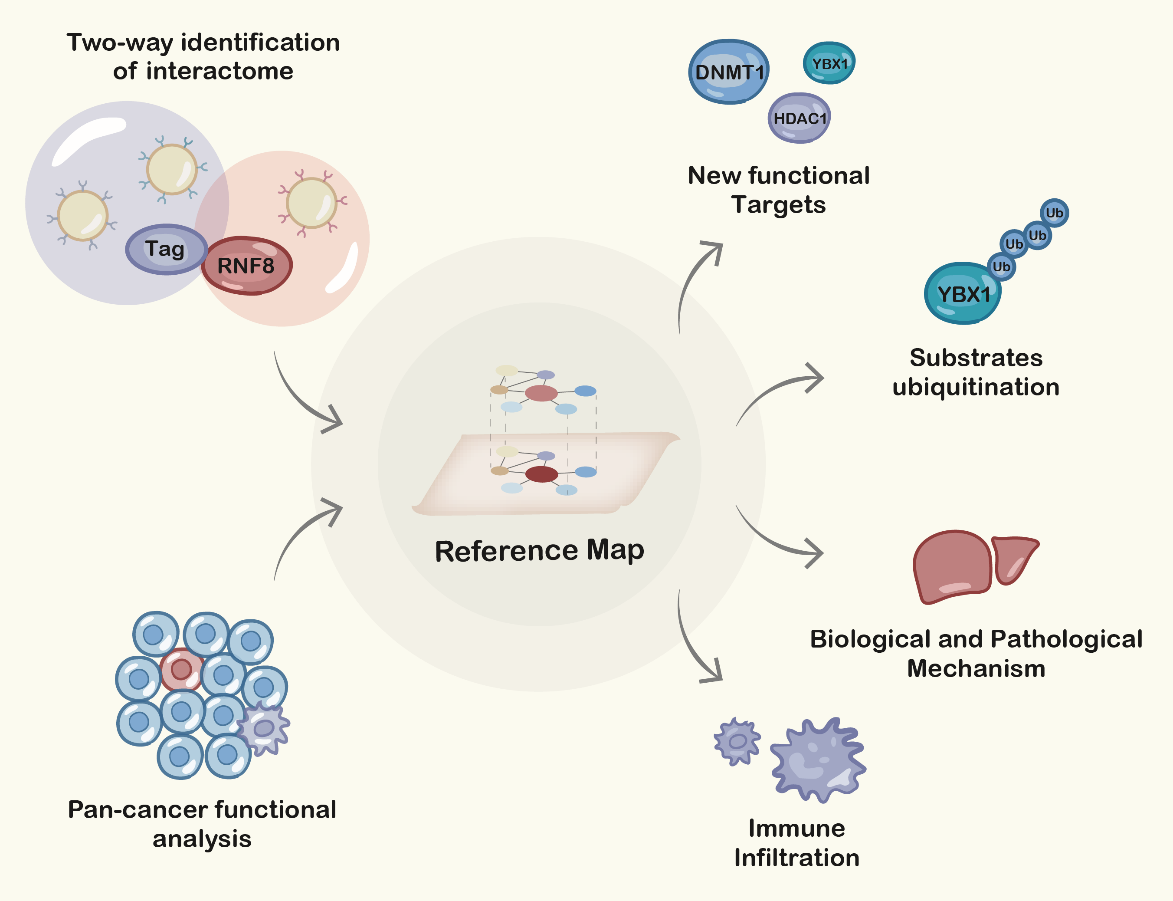
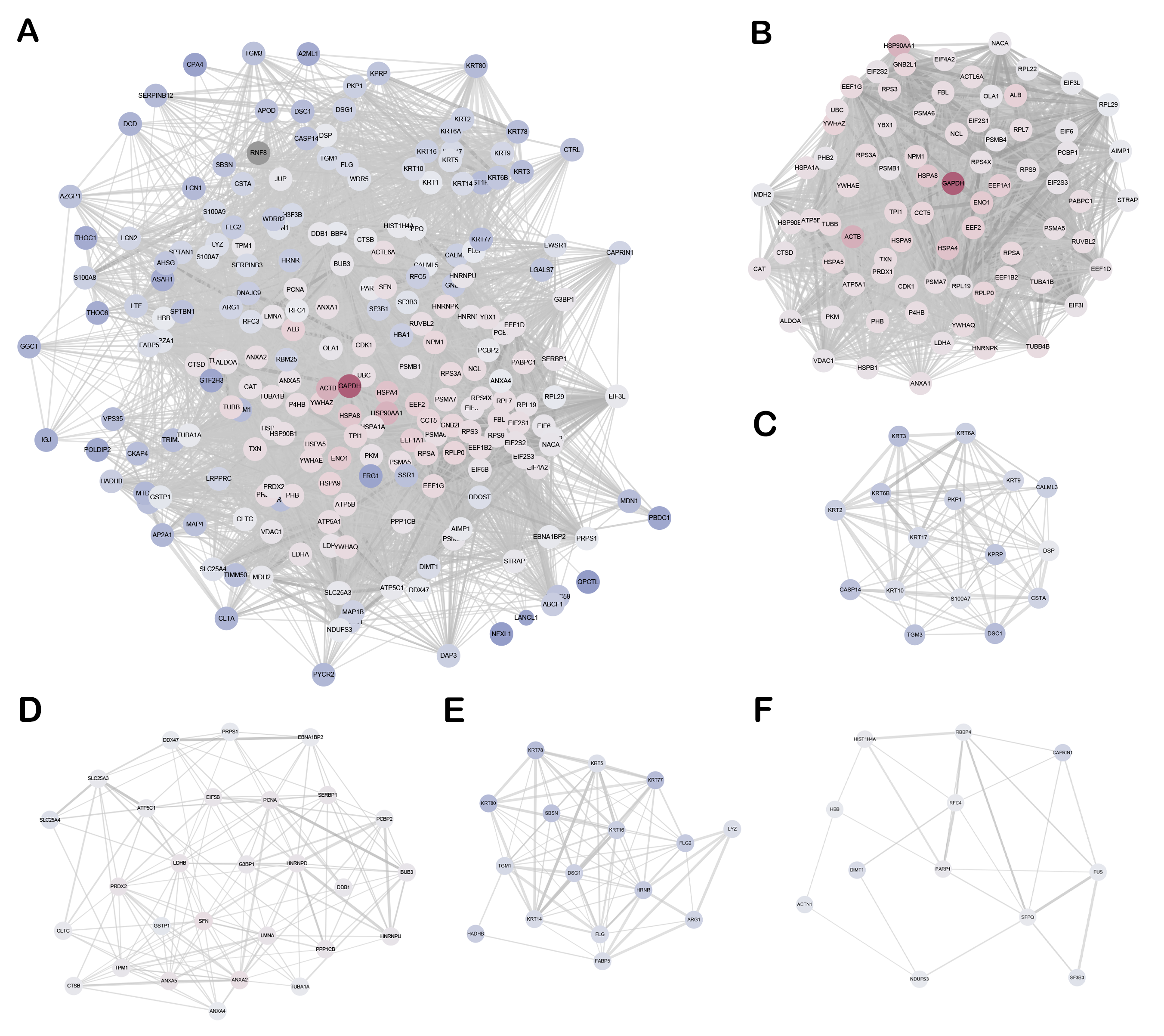
By integrating anti-RNF8-based IP with anti-Flag(+ anti-HA)-based IP, we identified RNF8 interactome with high specificity. our study provides a comprehensive reference map for RNF8 functions and reveals many new potential functions of RNF8 as well. In our results, some of the RNF8-targets pairs were demonstrated in previous reports, such as RNF8-UBC(42), etc., while most of the RNF8-targets pairs were not established previously. Given massive studies presented recently about RNF8 biological functions and potential therapeutic applications, the identification of RNF8 interactome surely would provide substantial evidence and instructions for RNF8-related mechanistic and functional studies.
As a proof of concept, we the interaction between RNF8-YBX1 to validate our interactome identification in vitro. YBX1 is a transcriptional factor that has been demonstrated to participle in the spliceosome, apoptosis, translation, cell proliferation, and tumor progression(43, 44). Recently, YBX1 has attracted a lot of attention. Gandhi et al showed that lincNMR regulated tumor cell proliferation through a YBX1-RRM2-TYMS-TK1 axis governing nucleotide metabolism(45). In addition, Goodarzi et al found that YBX1 stabilized pro-oncogenic transcript and enhanced cancer cell metastasis under hypoxia(46). In our study, we demonstrate the direct interaction between RNF8 and YBX1 and found that RNF8 promotes the ubiquitination of YBX1, which provides a novel molecular mechanism underlying YBX1-related biological processes. How RNF8-mediated YBX1 ubiquitination is involved in spliceosome and tumorigenesis needs to be explored in the future.
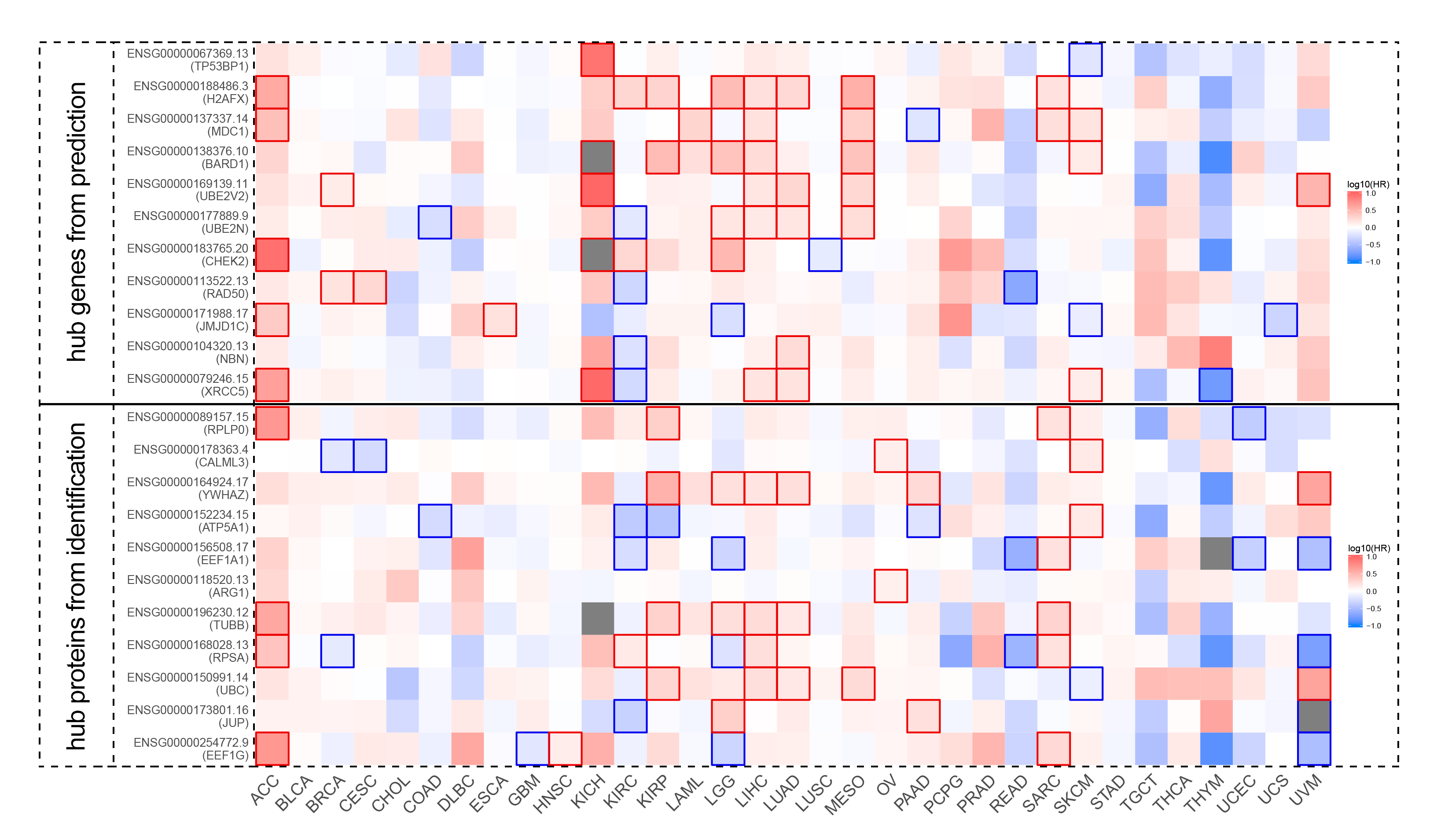
As shown in Table 1, our results for the first time identified the direct interaction between “Genome guardian” RNF8 and key genes in epigenetic regulation. Previous reports showed that RNF8-dependent ubiquitination of histone H2A during meiosis establishes active epigenetic modifications(47), however, how RNF8 regulates epigenetic modification remains largely unknown. As shown in Table 1, our results suggest that RNF8 might be a potential regulator for DNMT1, a key methyltransferase that maintains the methylation status after DNA synthesis and is associated with many important biological processes such as early embryo implantation and tumorigenesis, via direct interaction. In addition, HDCA1, a deacetylase which inactivates the expression of neuronal genes in non-nervous tissues and is implicated in axonal alteration and degeneration of the cell(48), is identified as the target of RNF8. Adam et al showed that RNF8-dependent polyubiquitination is required for the establishment of H3K27 acetylation. RNF8 was also reported to play a role in suppressing synapse formation(42) and neuron degeneration(49), but the clear mechanism underlying these physiological processes was poorly understood. Our findings might therefore provide new clues and evidence to these biological processes.
RNF8 was regarded as a DNA damage signal transducer. Upon DNA damage, the Mre11-Rad50-Nbs1 (MRN) complex senses the damage and recruit Ataxia telangiectasia mutated (ATM) to damage sites. ATM and ATR phosphorylated histone H2AX (referred to as γH2AX), and MCD1. RNF8 was then recruited to the DNA double-strand breaks site through FHA domain-mediated interaction with MDC1(50, 51) and stabilizes JMJD1C demethylase, demethylating MDC1 at K45, which promotes MDC1 association with RNF8(52). RNF8 then couples with Ubc13, DYRK2, L3MBTL2 to catalyze the formation of K63-linked polyubiquitin chains on many chromatin substrates, including histones H2A, H2AX, and H1(22, 53), which result in the recruitment of DNA repair proteins including 53BP1, BRCA1, and RAD51 to facilitate NHEJ or HR repair. RNF8 also regulates the abundance of the nonhomologous end-joining (NHEJ) repair protein KU80 and JMJ2A via catalyzing K48-linked polyubiquitination at sites of DNA damage. promotes efficient DSB damage repair by decreasing the pro-apoptosis activity of p53 through regulating Tip60 protein activity(22). Our results showed that RNF8 might regulate DNA repair via multiple targets such as PCNA, TP53, RAD51, CDK2, etc. Some of the targets are consistent with numerous reports, for example, Li demonstrated that PCNA is a target of RNF8 and monoubiquitinated by it. While most of the other targets and the function of RNF8-targets axes are yet to be explored.
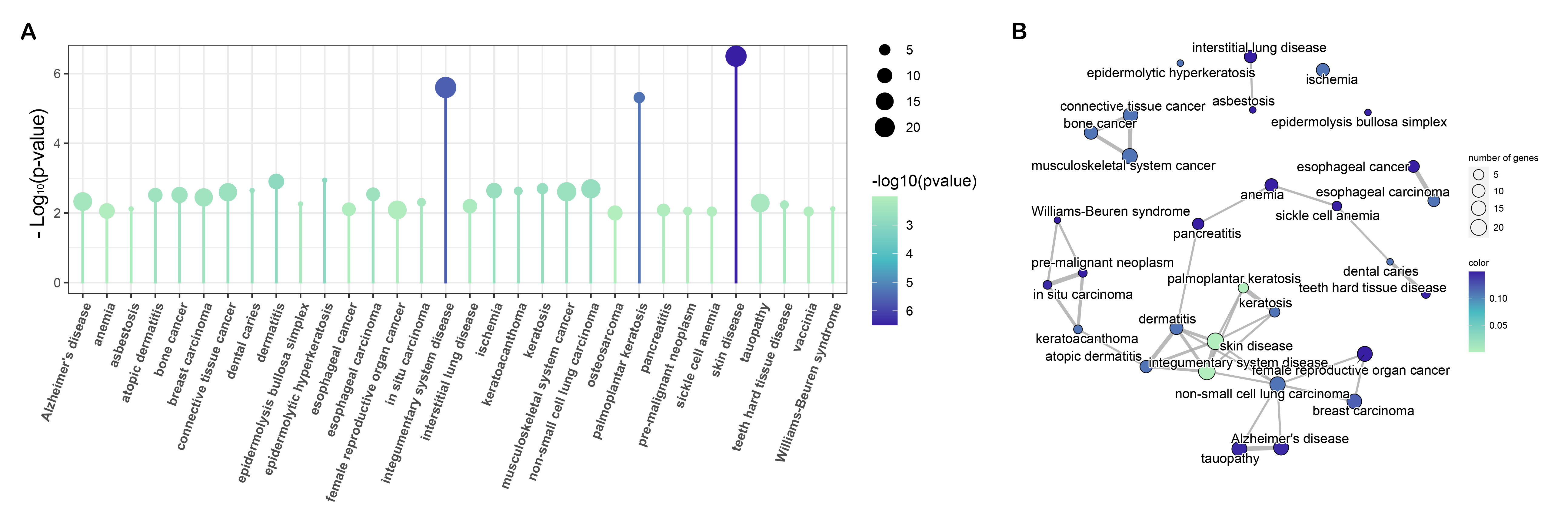
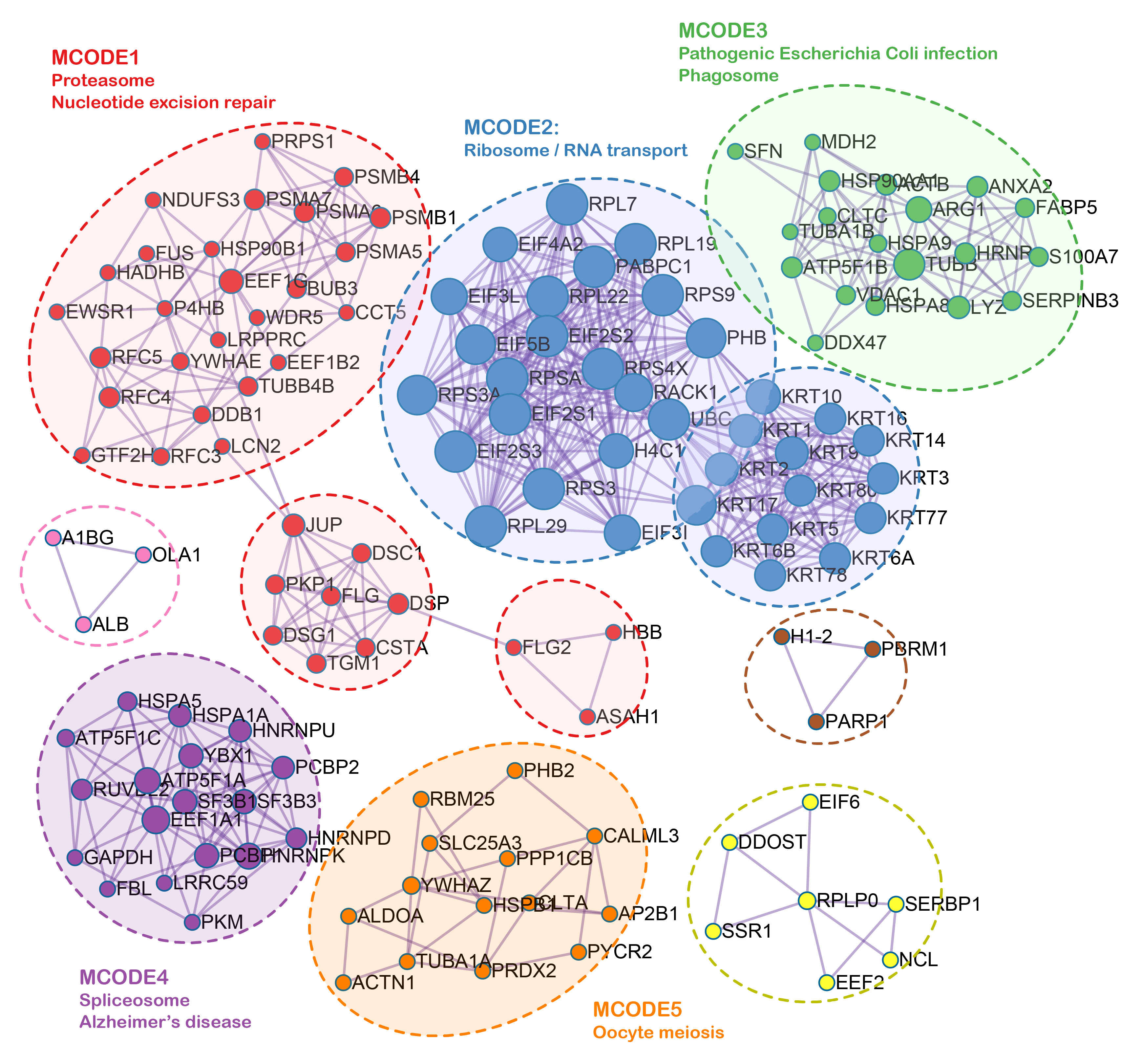
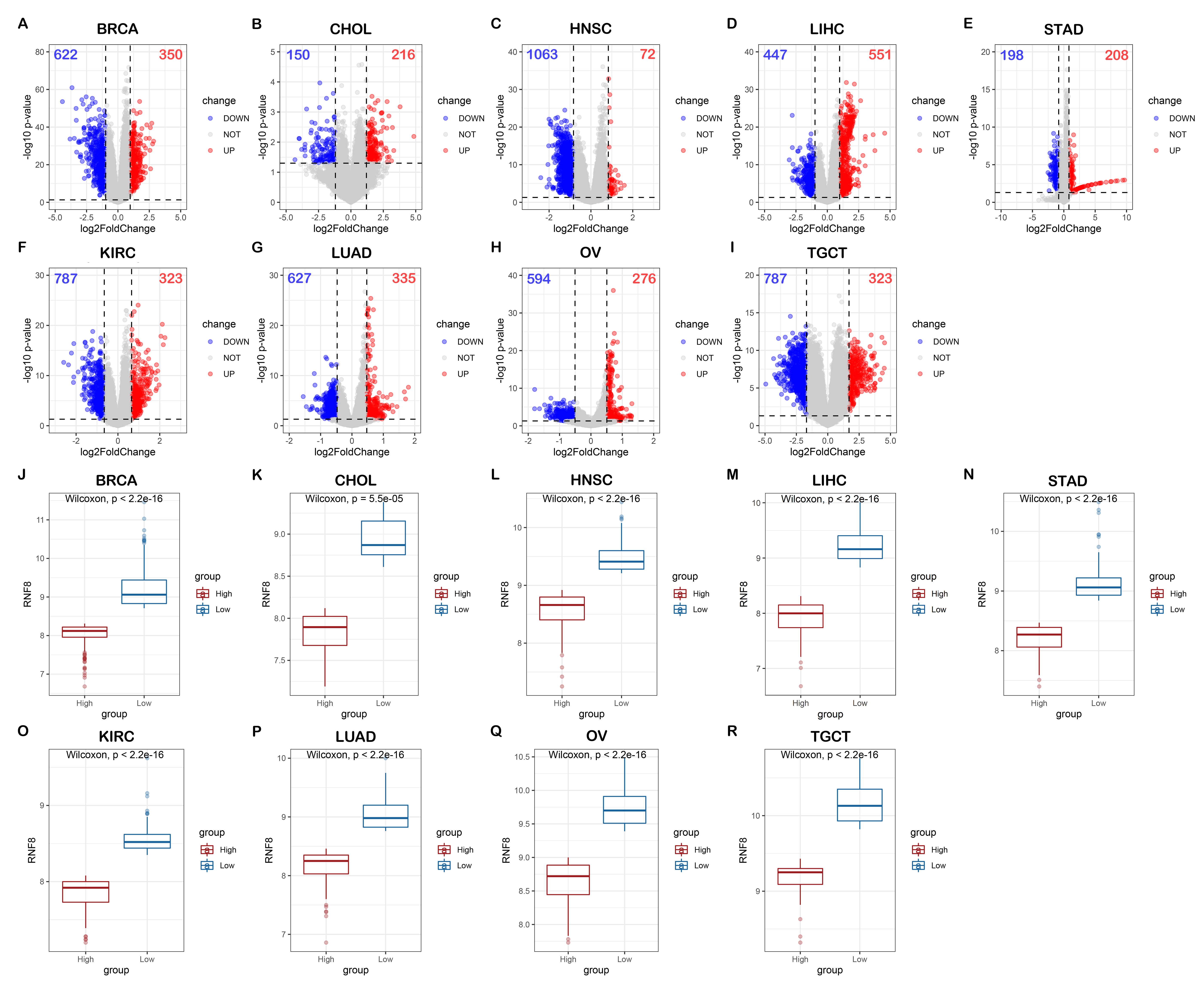
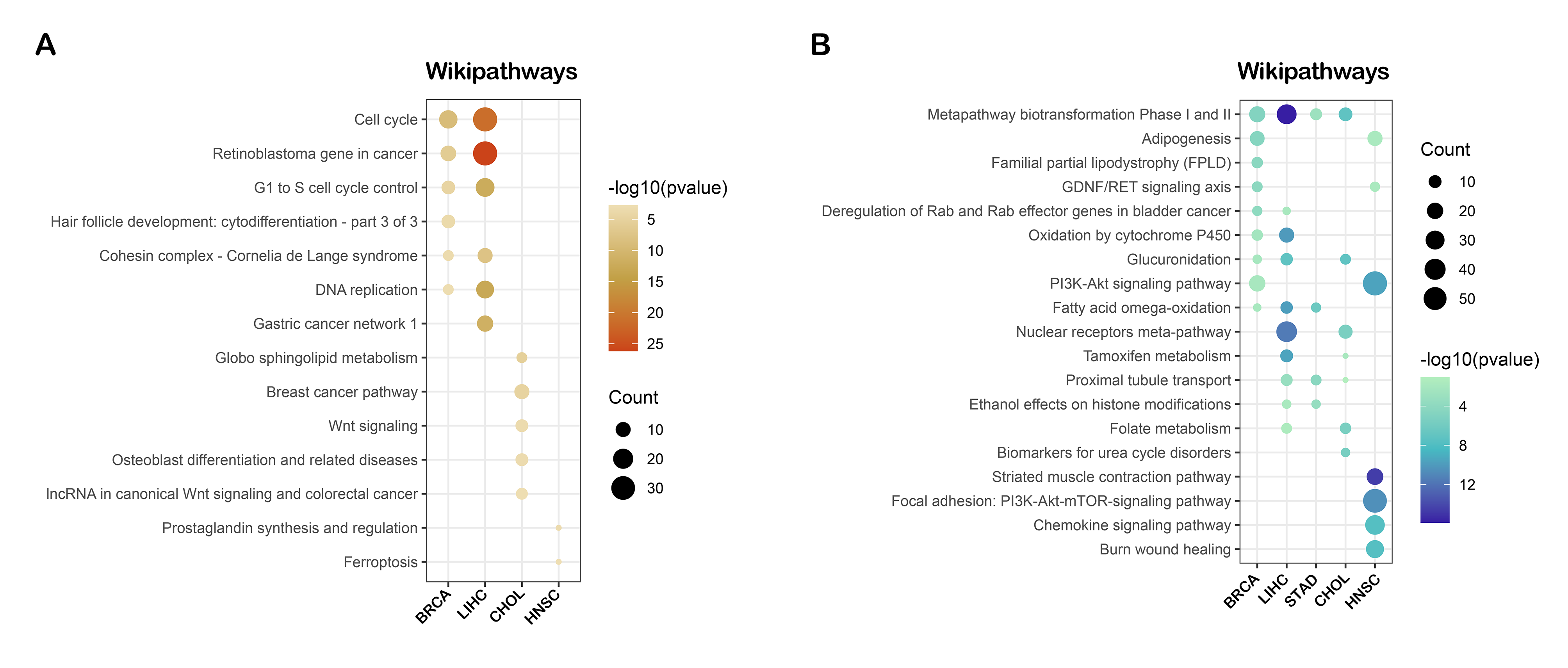
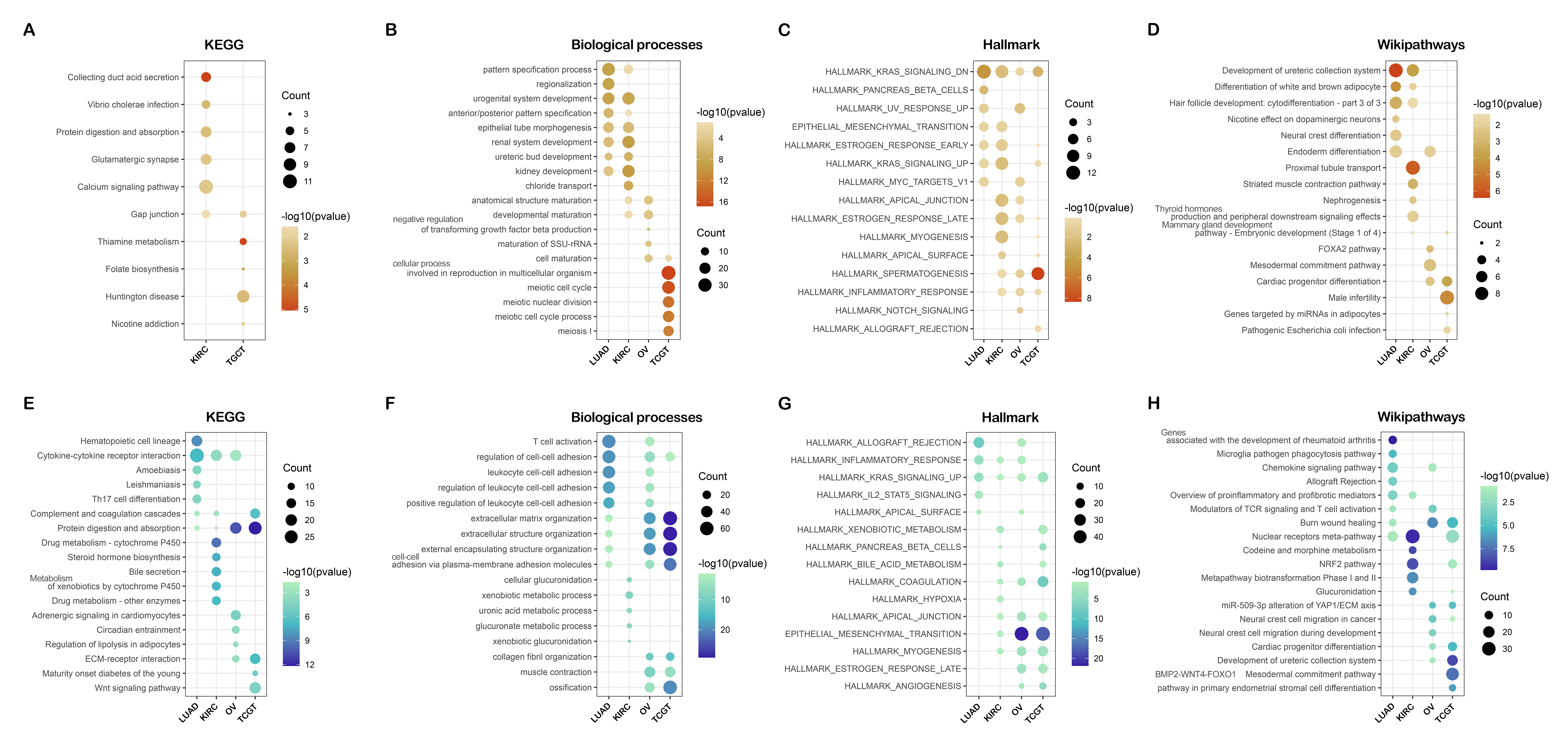
Though preliminary, we provide evidence demonstrating that RNF8 might be involved in many biological pathways. To reveal the common function of RNF8 in a larger scale. Integrated bioinformatics analysis and network analysis were therefore performed. Interestingly, in our KEGG enrichment analysis, the first thing we notice is that RNF8 was shown to be involved in many neurodegeneration diseases like Parkinson's disease, Alzheimer's disease, and amyotrophic lateral sclerosis (Fig. 2D), the relationship between RNF8 and these diseases might be a new area that needs to be further explored. as shown in Siwei’s study, RNF8 deficiency results in neurodegeneration in mice(49), but how RNF8 regulates the processes and how exactly RNF8 regulates neurodegenerative diseases still needs to be further explored and discussed based on identified interactome. Besides, the role of RNF8 in immunology-related processes such as neutrophil-mediated immunity, neutrophil activation, and whether these connections affect the role of RNF8 in cancer tissues has not been discussed and requires further exploration. The spliceosome is another item that is enriched in both MCODE and KEGG pathways. Many of our results show a strong connection between RNF8 and mRNA spliceosome, whether RNF8 regulates spliceosome, and whether the regulation is via YBX1 or other identified targets require further experimental validation.
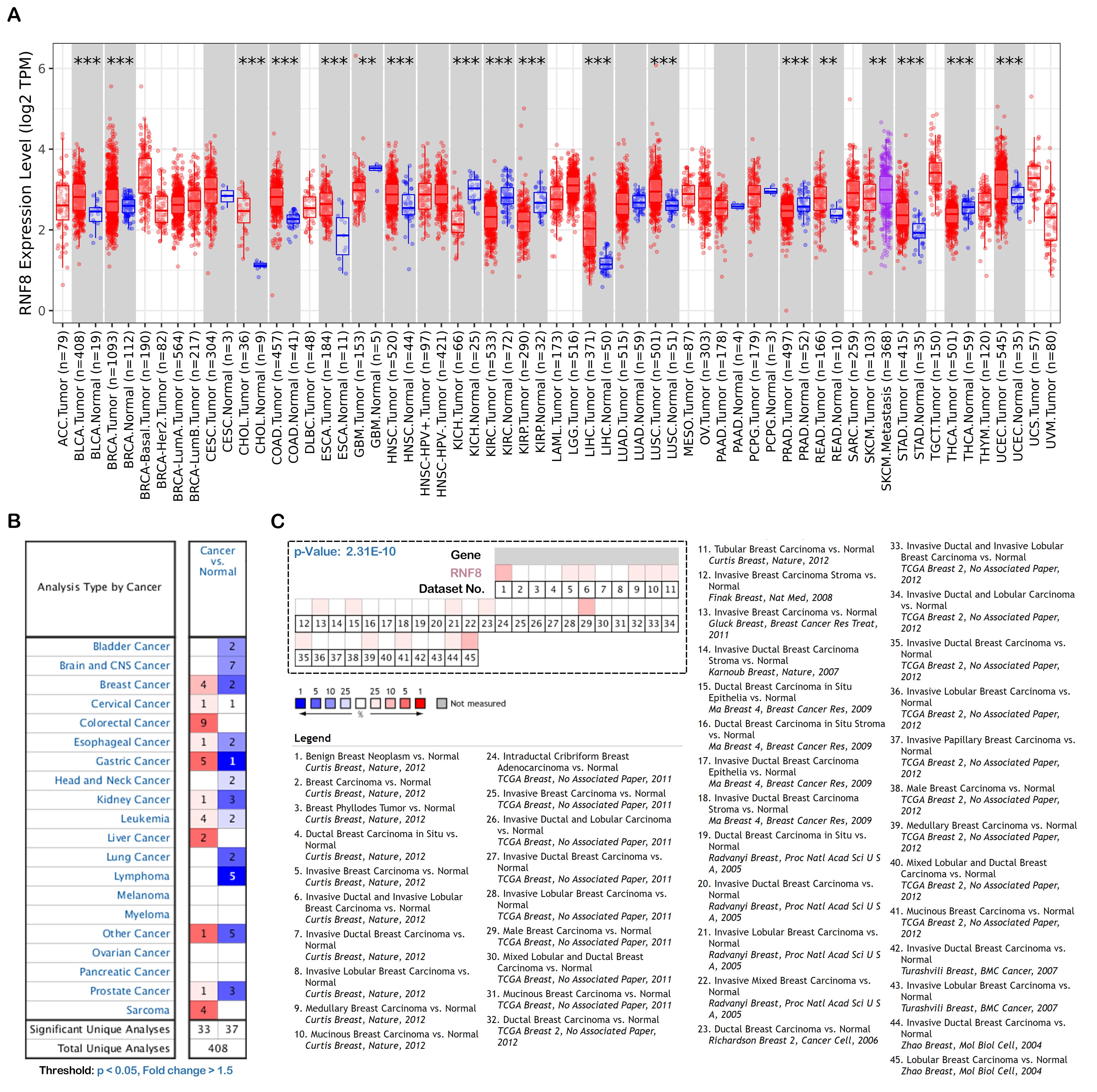
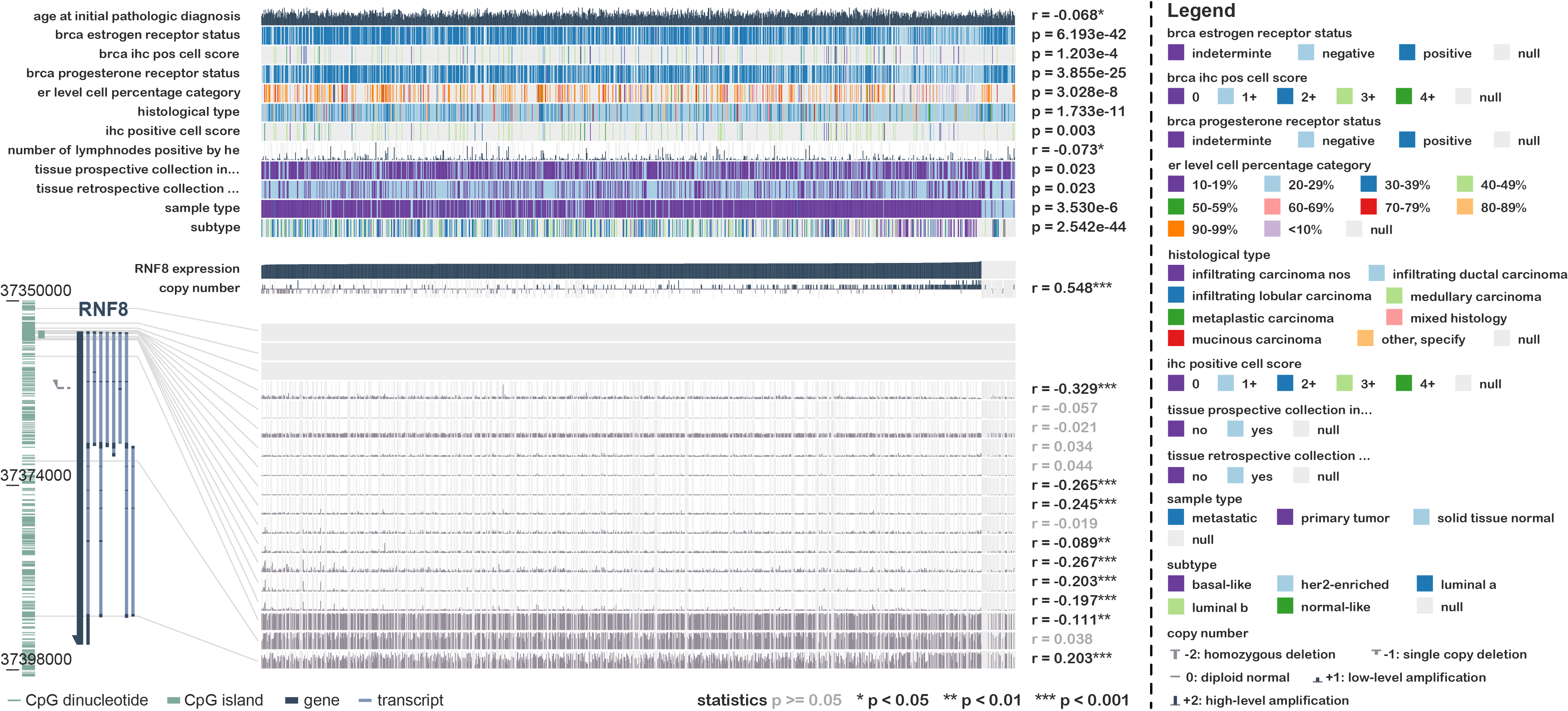
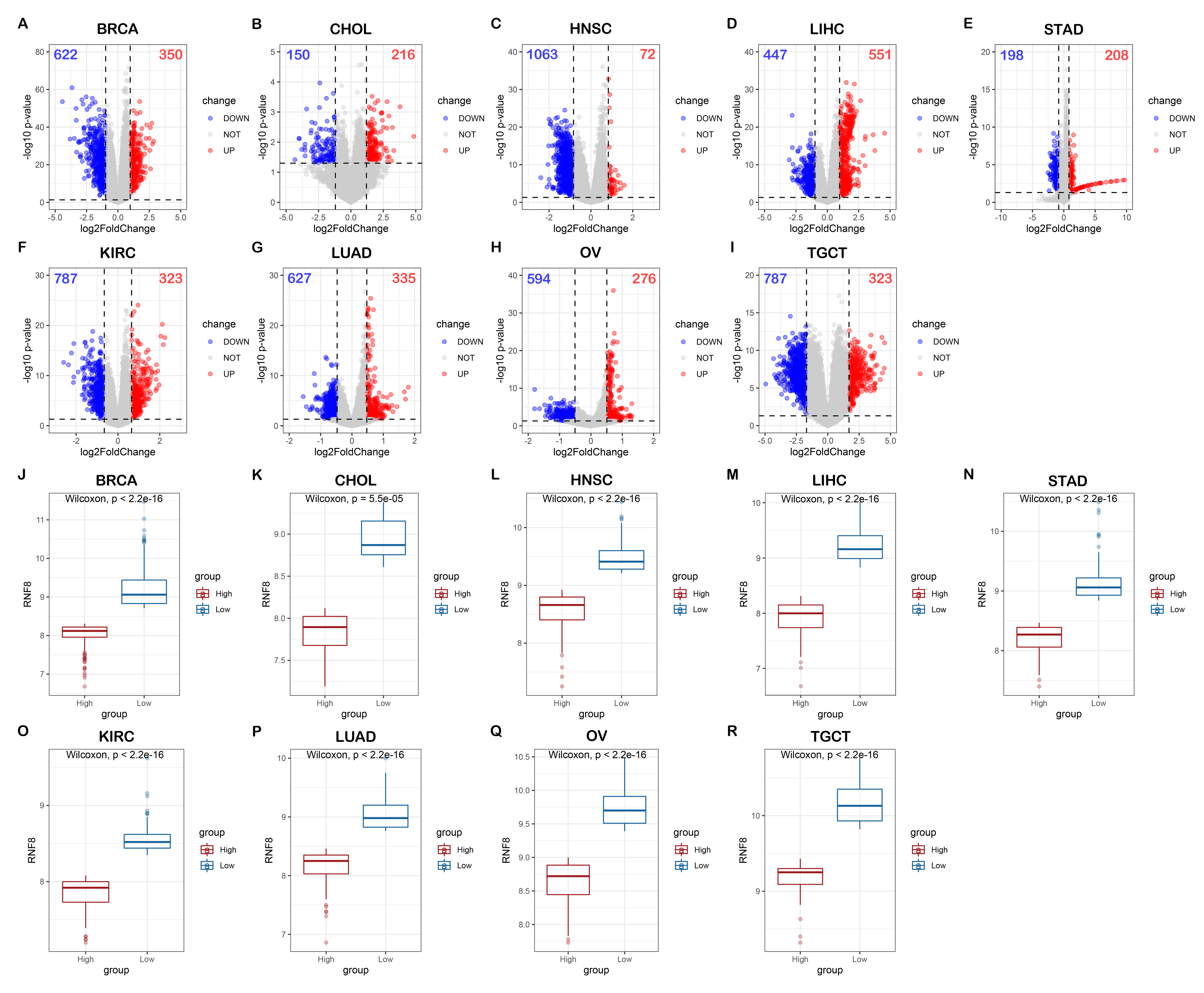
We noticed that the role of RNF8 in tumorigenesis, metastasis, and chemoresistance has been increasingly reported in many cancers recently. In breast cancer, RNF8 promotes cancer progression and metastasis through Twist activation, RNF8-mediated epithelial-mesenchymal transition is also regulated by multiple miRNAs such as miR-622, miR-214, which is consistent with our enrichment analysis results that RNF8-interactome might participate in cadherin binding and cell adhesion. With multiple new identified RNF8 targets, our identified interactome might be some alternative mechanism underlying these RNF8-regulated biological processes(4, 33, 54). In hepatocellular carcinoma, Trabid inhibits cancer growth and metastasis by cleaving RNF8-induced K63-linked ubiquitination of Twist(18). RNF8 also promotes tumorigenesis in lung cancer(16), silencing RNF8 sensitized bladder cancer to rediotherpy(55). Our results showed that RNF8 might participate in metabolic reprogramming in colon cancer and clear cell renal cell carcinoma. Ling et al showed that RNF8 can induce β-catenin-mediated c-Myc expression and thus promotes colon cancer proliferation, which is consistent with our findings. Also, our identification of RNF8-interacting ENO1, LDH1, PKM, PYCR, and MDH2 might provide other mechanisms underlying RNF8-regulated colon cancer progression.
In addition, our functional analysis about RNF8 in different cancers implied that RNF8 might be associated with the function of immune cells, these data quickly attracted our interest. As discussed above, lots of efforts have been made to discover how RNF8 regulate cancer progression, metastasis, and prognosis, while in the view of tumor micro-environment, whether RNF8 influence the physiological status of tumor-infiltrating immune cells, a class of immune cells playing critical roles in cancer progression and closely related to clinical outcomes, are largely unknown. By analyzing bulk RNA-seq data and scRNA-seq data, our results showed a direct correlation between RNF8 expression and immune cells such as monocytes, macrophages, etc. in various cancers. These findings might provide new insight into the RNF8-regulated tumor-associated biological processes and mechanisms.
In summary, our identification of the RNF8 interactome revealed numerous new targets of RNF8. Based on these identified targets and integrated bioinformatic analysis, we systematically revealed the potential functions of RNF8 at the protein-protein interaction level and pathways level. We believe our work will provide a unique framework for researchers and clinicians who seek to better explore or understand RNF8-regulated biological functions, as well as its clinical applications.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}