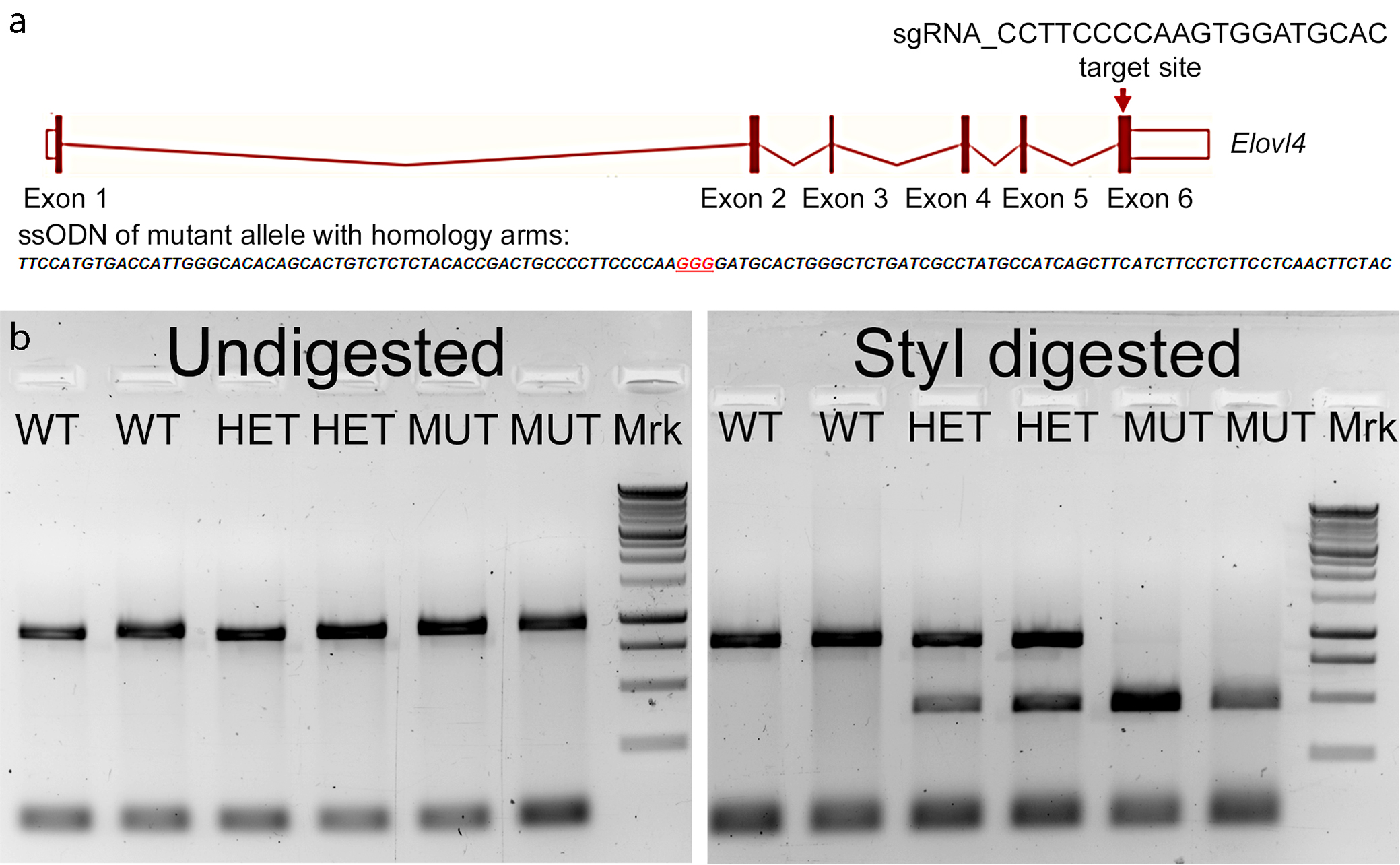
Generation of the SCA34 rat model
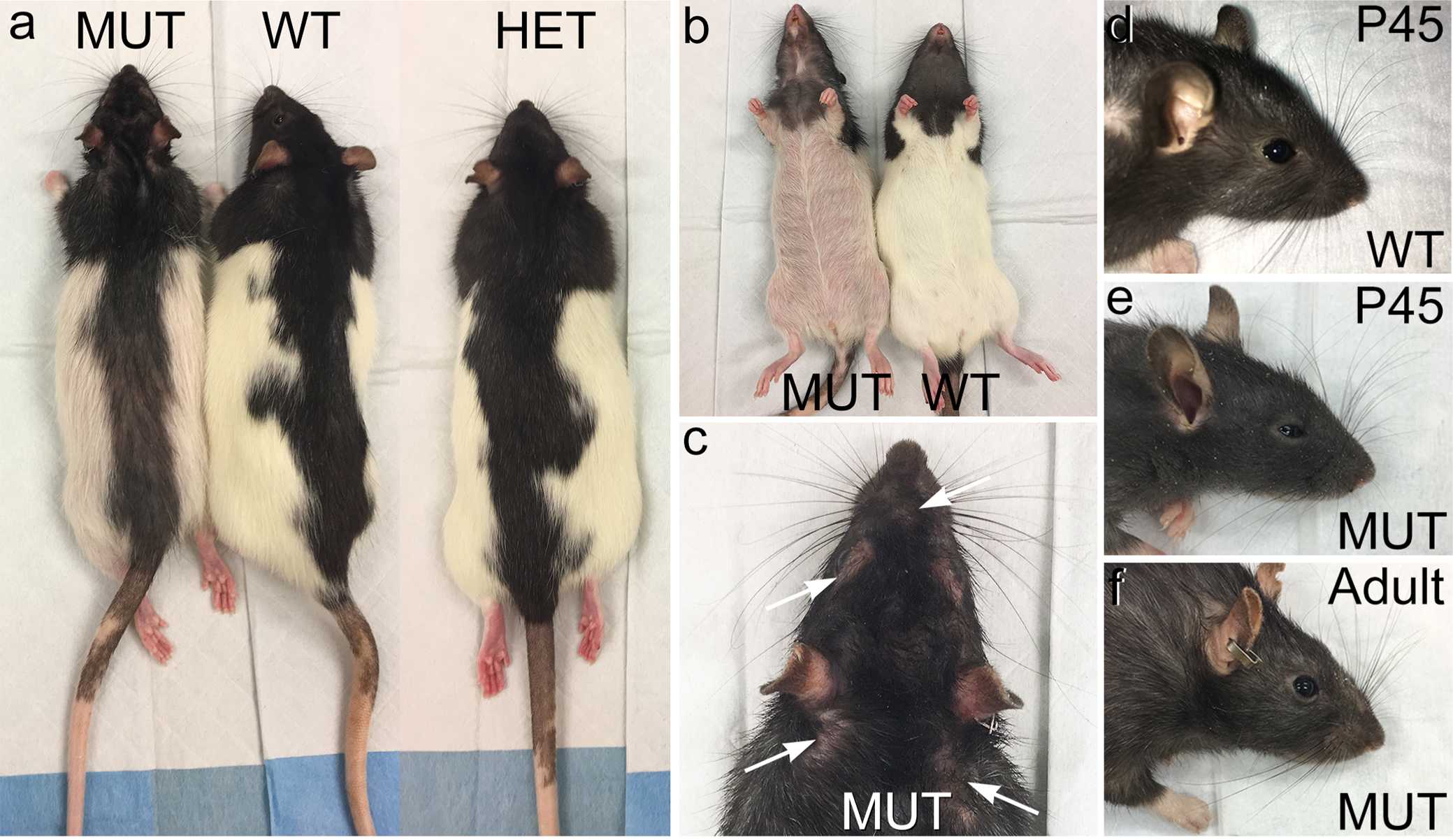
We used the CRISPR-Cas9 gene editing approach to knock in the c.736T>G (p.W246G) mutation that causes human SCA34 [16] into exon 6 of the rat Elovl4 gene (Ensembl: ENSRNOG00000009773) to generate heterozygous Long-Evans F0 founder rats [Fig. 1A. See Agbaga et al., 2020 for a more detailed description [34]. The characterization of the model is reproduced from (34) in Supplemental Figures 1-6]. The F0 founder rats were bred to wild type (WT) Long-Evans rats to establish heterozygote breeders. For consistency and clarity, this mutation is referred to as the "W246G ELOVL4 mutation." Editing of a single copy of the WT Elovl4 allele to the c.736T>G (p.W246G) mutant allele was confirmed by genotyping and by Sanger and whole genome DNA sequencing, which did not find any aberrant indels, insertions, or off-target editing effects [34]. Heterozygous W246G ELOVL4 knock-in rats were fertile and passed the 736T>G allele to their offspring to produce heterozygote (HET) and homozygote (MUT) offspring. MUT rats were viable and fertile, in contrast to mice with Elovl4 knockout or homozygous expression of STGD3 alleles, which die from dehydration shortly after birth due to loss of the skin water barrier [30-33]. Phenotypically, MUT rats presented with EKV characterized by reddened skin and patchy loss of hair [34], consistent with the finding of EKV in some human SCA34 patients [14,15]. Age-matched female rats showed no differences in weight across genotypes from 2 to 6 months of age. In contrast, male rats exhibited a sex-related difference in weight, with male MUT rats being significantly smaller than age-matched male WT and HET rats from 2 to 6 months of age (Fig. 1B). Immunoblotting showed no differences in ELOVL4 levels in the WT, HET, and MUT cerebellum (Fig. 1C-D) using a well-characterized antibody that recognizes WT and full length MUT ELOVL4 [1]. Finally, assessment of the effect of the W246G ELOVL4 mutation on VLC-SFA and VLC-PUFA synthesis showed that the mutation significantly reduced, but did not eliminate, VLC-SFA production in the skin (remaining 28:0 + 30:0 levels around 35% compared to WT), and had no significant effect on VLC-PUFA synthesis in the retina (34; Supplemental Figs. 5 and 6).
The W246G ELOVL4 mutation did not cause overt impairment of motor function, as male and female HET and MUT rats could walk, rear on their hind legs, and breed, consistent with the relatively late onset and slow progression of SCA34 in humans with the W246G ELOVL4 mutation. To better assess effects on motor performance, cohorts of female WT, HET, and MUT rats were tested at 2, 3, 4, and 6 months of age using the rotarod assay. The rotarod test revealed significant motor impairment in female HET and MUT rats compared to WT rats at all ages examined (Fig. 2), indicating that the deficits observed at 2 months were not transient. Interestingly, HET and MUT female rats performed equally poorly in the test. These studies indicate that the W246G ELOVL4 mutation caused persistent motor impairment in HET and MUT rats starting from early ages. Male rats of all three genotypes performed unreliably when tested on the rotarod task at 2 to 6 months of age (data not shown).
The W246G ELOVL4 mutation does not disrupt ELOVL4 distribution or cerebellar cytoarchitecture out to at least six months of age.
Cerebellar atrophy is a common feature of SCA34 [14,16] and loss of Purkinje cells in particular is a prominent feature of many forms of SCA [43]. To assess whether the W246G ELOVL4 mutation caused neurodegeneration or disrupted cerebellar cytoarchitecture, we conducted a detailed morphological assessment of the cerebellar cortex. Gross cerebellar structure was not affected by the W246G ELOVL4 mutation (Fig. 3A-I). Lobule organization was similar in WT, HET, and MUT rats through at least 6 months of age. The cerebellar cortex showed the normal three-layered structure comprised of a thick, outer molecular layer housing interneurons and synapses, a well-organized monolayer of Purkinje cells, and a densely packed granule cell layer adjacent to the deep white matter of the arbor vitae (Fig. 3J-L). Analysis of cortical layer thickness in lobules 3, 5, and 10 showed no thinning of the molecular or granule cell layers at 3 or 6 months (Fig. 3M-T). A difference in the thickness of the Purkinje cell layer was noted at 6 months of age in lobule 3, but the linear density of Purkinje cells showed no differences across ages or genotypes (Fig. 3U-V), indicating that there was no substantial loss of Purkinje cells over this time frame.
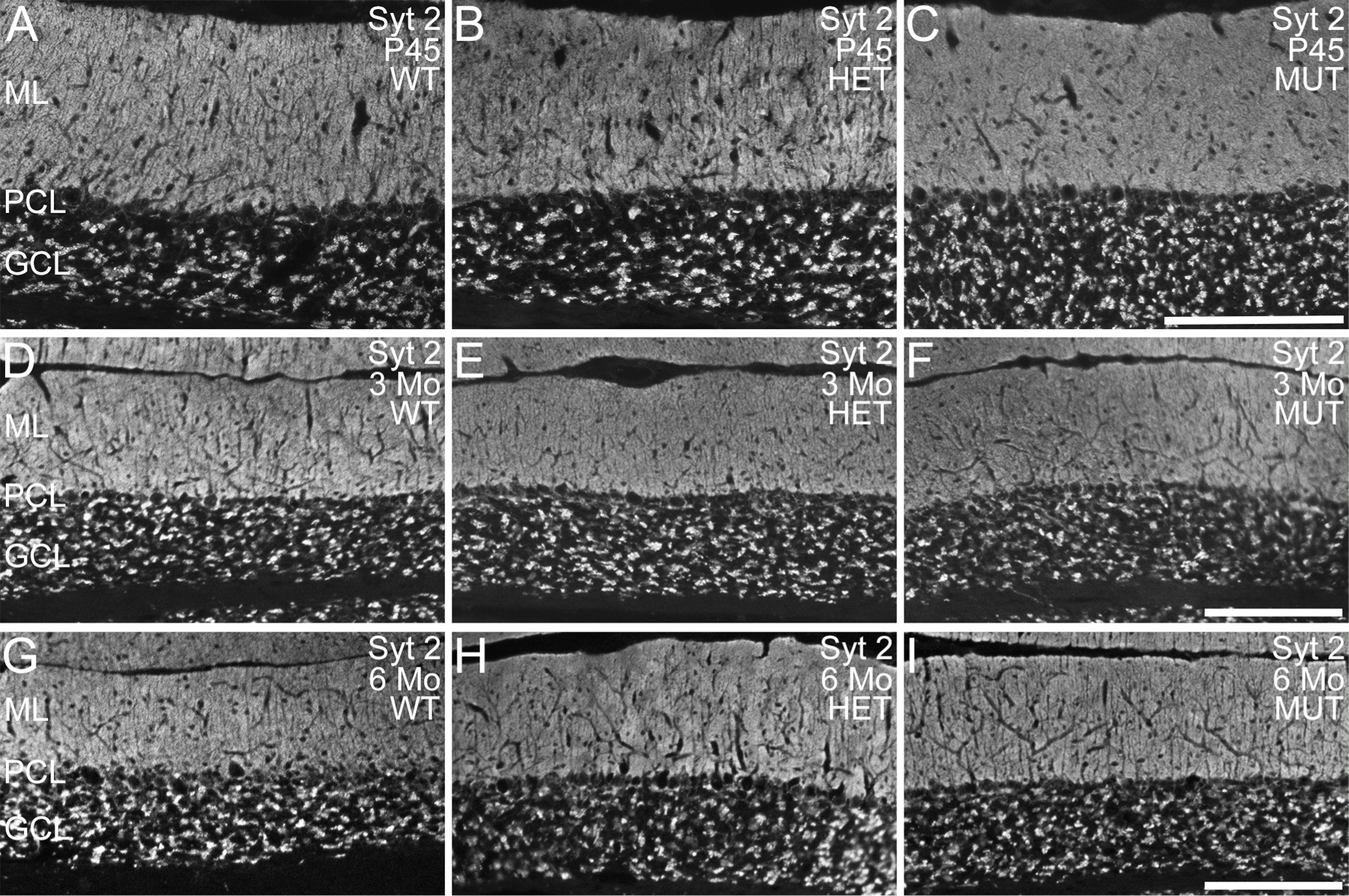
To further assess cerebellar integrity, immunolabeling for ELOVL4 and a variety of well-known cell- and synapse-specific markers was performed (Fig. 4). Notably, as the ELOVL4 antibody recognizes both wild type and mutant protein, we were able to detect the distribution of WT and W246G mutant ELOVL4 protein using the same antibody (see Fig. 1). The W246G ELOVL4 mutation did not alter ELOVL4 distribution in the cerebellum. ELOVL4 was present in granule cells, inhibitory interneurons in the molecular layer, and Purkinje cells in the cerebellum of rats of all genotypes (Fig. 4A-C), similar to ELOVL4 distribution in the mouse cerebellum [37]. Purkinje and granule cells labeled for calbindin and NeuN, respectively, showed normal morphology and distribution in WT, HET, and MUT rats at all ages examined (Fig. 4D-I; Supplemental Figs. 7,8). Labeling for VGluT1, a marker for mossy and parallel fiber synapses, and synaptotagmin 2, a marker for GABAergic synapses, both showed numerous synapses throughout the molecular layer and in the synaptic glomeruli of the granule cell layer in all genotypes at all ages, as appropriate (Fig. 4J-O; Supplemental Figs. 9,10). Labeling for VGluT2, a specific marker for climbing fiber synapses onto Purkinje cells, confirmed that climbing fiber synapses were present in the molecular layer of WT, HET, and MUT cerebellar cortex, as expected (Fig. 5). The distribution of mGluR1 [44], which is critical to Purkinje cell synaptic plasticity [45], also was unaffected by the W246G ELOVL4 mutation (Fig. 5). Labeling for astrocytes and microglia showed the normal distribution and morphology of these cell populations across genotypes of all ages, with no sign of overt neuroinflammation (Supplemental Figs. 11, 12).
To further assess the effects of the W246G ELOVL4 mutation at the synaptic level, western blot analysis of several important synaptic proteins associated with glutamatergic and GABAergic synapses was performed (Fig. 6). We examined the following proteins: excitatory presynaptic markers: VGluT1, VGluT2; AMPA glutamate receptors: GluA1, GluA2 and the postsynaptic density protein PSD-95; and inhibitory synaptic markers: Syt2, GAD65, and GAD67. No significant differences were present among WT, HET, and MUT cerebellum for any of the glutamatergic and GABAergic synaptic proteins examined.
Together, these data indicate that the motor deficits observed in HET and MUT rats did not arise from any gross disruption of the cellular or synaptic organization of the cerebellum or neurodegeneration.
The W246G ELOVL4 mutation alters neuronal network communication in the cerebellar cortex.
Cerebellar cortical function is essential for motor learning and it is thought to involve fine tuning of the extensive PF-Purkinje cell synaptic network in coordination with the “error reporting” of CF-Purkinje cell input [46]. To better understand the origins of the motor impairment associated with the W246G ELOVL4 mutation, we assessed neuronal network properties in the cerebellum using multi-electrode array electrophysiological recording from cerebellar slices [47] prepared from WT and MUT rats (Fig. 7, see Methods for details). Briefly, an array of 61 electrodes arranged hexagonally at 70 µm intervals was carefully positioned under the slice to cover all cerebellar cortical layers. The overall spontaneous network activity was recorded for 10 min under resting conditions [3]. Assessing entire ten minute-long recordings and plotting the events per second for all 61 electrodes revealed two major differences in network activity in WT and MUT slices (Fig. 7A). First, slices generally showed either very low activity (less than average of 1 event per sec, upper panels) or high activity (lower panels on Fig. 7A). There was no significant difference between WT and MUT slices in the distribution of low to high activity (20/9 for WT and 15/10 for MUT, Chi square value is 0.4732, p=0.491511). However, there was a striking failure of PF synchronization in MUT slices compared to the almost uniform synchronization of spontaneous activity in WT slices recorded on all electrodes simultaneously (Fig. 7A, lower two panels). Interestingly, in many MUT slices, we detected robust, persistent activity (Fig. 7A, lower two panels). Next, we analyzed spike frequency, amplitude, and interspike interval (ISI) from these recordings. Although average spike frequency did not differ in the cerebellar cortex between WT and MUT slices (Fig. 7B left panel), further analysis revealed an increased average frequency of positive spikes (Fig. 7B middle panel) and a reduced frequency of negative spikes in the MUT cerebellum (Fig. 7Bright panel). In addition, the amplitude of network events was significantly decreased in MUT cerebellar slices compared to WT (Fig. 7C, negative spike traces and positive spike traces; and Fig. 7D). The overall distribution of events as shown by the ISI histogram was similar in WT and MUT slices (Fig. 7E, p=0.219, Wilcoxon ranked test), with both showing peak activity at two frequencies of 0.5 Hz and 0.26 Hz. We noticed more activity at the highest frequencies in the WT slices and selectively analyzed the 1Hz or higher activity signals (Fig. 7F). Indeed, MUT cerebellar slices had a marked reduction in activity above 20 Hz (ISI <0.05 s; Fig. 7F; p<0.0001, Mann-Whitney test). Next, to determine if PF activity was altered under resting conditions, we analyzed the spontaneous activity from the same electrodes that were used later to record PF LTP. Activity in MUT PF differed from WT PF: the difference was mostly in the high >20Hz frequencies, but interestingly, the 0.6-4 Hz activity corresponding to delta band was also reduced (Fig. 7G; p<0.0001, Mann-Whitney test). These observations from the spontaneous network activity indicates that W246G ELOVL4 mutation alters and desynchronizes neuronal network communication in the cerebellar cortex.
The W246G ELOVL4 mutation severely impairs synaptic plasticity at PF and CF inputs to Purkinje Cells.
Differences in neuronal network activity between WT and MUT cerebellum as well as the motor learning deficits of HET and MUT rats in the absence of neurodegeneration are consistent with the notion that synaptic function is impaired by the W246G ELOVL4 mutation. Therefore, we tested whether the W246G ELOVL4 mutation affected neurotransmission and long-term synaptic plasticity in the cerebellum. Long-term potentiation (LTP) and long-term depression (LTD) are hours-long increases and decreases, respectively, in synaptic strength in response to strong synaptic stimulation, and are important to motor learning. ELOVL4 is highly expressed in cerebellar granule cells (Fig. 4) and cells in the inferior olivary complex [37], and its VLC-SFA products that are incorporated into sphingolipids are known to modulate synaptic release in hippocampus [3]. To assess the functional effects of the W246G ELOVL4 mutation on long-term synaptic plasticity, we tested the function of PF and CF inputs to Purkinje cells in cerebellar slices prepared from 3-4 month old WT and MUT rats.
The most numerous excitatory synaptic inputs to Purkinje cells are from PFs, which drive simple spikes in Purkinje cells [48]. To assess the function of PF-Purkinje cell synapses, acute ex vivo cerebellar slices from WT or MUT rats were stimulated using one electrode located in the molecular layer to stimulate PFs. The field potentials of excitatory post-synaptic potential (fEPSP) responses in the molecular layer were recorded every 30 sec using a MED64 multi-electrode array system (MEA). A stimulation paradigm of 1 Hz for 10 min was used to induce PF LTP [modified from [49-52]]. This stimulation paradigm does not require CF co-stimulation and indeed induced robust LTP in slices from WT but not MUT rats (Fig. 8). As expected, WT slices showed a significant increase in the slope of PF fEPSP one hour after stimulation. However, no increase in the PF fEPSP slope was present in slices from MUT rats, indicating severe impairment of LTP at PF-Purkinje cell synapses [Fig. 8A, upper panel, WT = 145.4 ± 9.2% normalized slope, n=14 slices from 6 rats; MUT = 96.2 ± 3.8%, n=16 slices from 6 rats; Mean ± SEM; *** p < 0.001, two-way repeated measures ANOVA with Bonferroni post-hoc test (RM ANOVA)]. We also measured the basic physiological properties of the LTP at PF-Purkinje cell synapses by recording the characteristics of synaptic responses to a range of increasing stimuli (5-100 μA) before and after recording LTP. This “Input–Output” test allowed us to assess general physiological properties of PF – Purkinje cell synapses and to evaluate how LTP affects them. We found that, at stimulus intensity greater than 60 μA, MUT slices showed a significantly smaller fEPSP amplitude than WT slices before LTP (Fig. 8B; lower left panel, * p < 0.05, two-way RM ANOVA). Furthermore, after LTP induction, while the response for WT slices increased significantly (Fig. 8B; upper left panel, *** p < 0.001, two-way RM ANOVA), MUT slices showed no change in response (Fig. 8B; upper right panel, ns, two-way RM ANOVA), indicating that MUT animals had severely impaired synaptic plasticity. Accordingly, the difference between WT and MUT slices in synaptic response to step-wise stimuli became even more pronounced after LTP was induced (Fig. 8B; lower right panel, ** p < 0.01, two-way RM ANOVA).
As the baseline synaptic strength differed between WT and MUT PF fiber recordings, we next scrutinized the synaptic responses during the 1 Hz stimulus trains. We found that the applied low intensity stimuli evoked similar responses from both genotypes (Fig. 8D), as the average amplitude of the first response was 395 ± 81.6 µV for WT and 344.3 ± 100 µV for MUT (p=0.66, Mann-Whitney test). During the stimulus train WT responses increased significantly, but MUT responses remained stable (Fig. 8C, ** p < 0.01, two-way ANOVA, last 100 sec). This also was reflected in the ratio of average amplitude of the last five responses divided by the average of the first five responses (Fig. 8E), which increased during the 1 Hz stimulation only in WT slices (*** p < 0.001, Mann Whitney test). Taken together, these observations support the conclusion that PF synapses with the W246G ELOVL4 mutation are selectively defective in LTP, although they respond perfectly well to low-intensity single stimulation.
A second key excitatory synaptic input to the Purkinje cells comes from the CFs, which drive complex spikes in Purkinje cells and are critical to motor learning [48]. The CF-Purkinje cell synapse has a high release probability [53] and shows pronounced LTD in response to tetanic stimulation. To assess the function of CF-Purkinje cell synapses, slices from WT or MUT rats were given tetanic stimulation to activate CFs at low frequency (1 Hz for 10 min) and CF fEPSP responses were recorded at the boundary of Purkinje cell and molecular layers every 30 sec using the MEA system [modified from [49]]. This stimulation paradigm induced strong LTD in slices from WT rats as expected, but did not induce LTD in slices from MUT rats (Fig. 9A). The level of depression was significantly higher in slices from WT rats compared to slices from MUT rats one hour after induction, indicating severe impairment in LTD in the presence of the ELOVL4 W246G mutation (WT = 60.5 ± 4.6% normalized slope, n=10 recordings from 5 rats; MUT = 95.98 ± 3.4%, n=14 recordings from 6 rats; Mean ± SEM; *** p < 0.001, two-way RM ANOVA). We also recorded the Input–Output characteristics of synaptic responses to increasing stimuli (5-100 μA) before and after recording LTD to measure physiological properties of CF to PC synapses. We found that, at stimulus intensity greater than 50 μA, MUT slices showed a significantly smaller fEPSP amplitude than WT slices before LTD (Fig. 9B; lower left panel, *** p < 0.001, two-way RM ANOVA). However, after LTD induction, while the response for WT slices decreased significantly (Fig. 9B; upper left panel, *, p < 0.05, two-way RM ANOVA), MUT slices showed no change in response (Fig. 9B; upper right panel, ns, p > 0.05, two-way RM ANOVA), indicating that MUT animals had impaired LTD. Moreover, after LTD induction, we found that WT and MUT slices still responded differently to strong stimuli (>50 μA), with MUT slices showing a significantly smaller amplitude than WT slices (Fig. 9B; lower right panel, *, p < 0.05, two-way RM ANOVA). Finally, we analyzed the synaptic responses from CF during the 1 Hz stimulation (Fig. 9C-E), as described above for PF recordings. No significant difference was present between WT and MUT slices during the 600 pulses at 1Hz.
These experiments together indicate that the W246G ELOVL4 mutation affected both LTP at PF-Purkinje cell synapses and LTD at CF-Purkinje cell synapses, which are essential to normal cerebellar learning.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}