Thermodynamically, lactic acid and ethanol allow the best energy recovery among all common metabolites for methane production. The main objective of this work was to favor the fermentation of OFMSW for better lactic acid and ethanol production. First, OFMSW fermented with three different inocula: Naturally occurring microorganisms (NOM), yeast (Saccharomyces cerevisiae), and UASB sludge. Second, methanization of digestates from the fermentations. OFMSW fermentation with NOM and UASB sludge + NOM mainly produces lactic acid; yeast fermentation mainly produces ethanol, lactic acid, and volatile fatty acids. Methane production increased with decreasing substrate concentration, and higher methane production was obtained with the digestate from NOM fermentation. Every one of the fermented digestates produced more methane than unfermented OFMSW. Higher concentrations of propionic and butyric acids were observed for higher digestate concentrations from yeast and NOM after 30-day methanization. Methane produced from unfermented OFMSW was not inhibited as propionic, and butyric acids concentrations were the lowest of all four cases.
Research Article
OFMSW fermentation with different inocula and its effects on methane production
https://doi.org/10.21203/rs.3.rs-1463070/v1
This work is licensed under a CC BY 4.0 License
You are reading this latest preprint version
Alcoholic fermentation
lactic fermentation
anaerobic digestion
methane production
This research proposes a simple fermentation method to produce the best eligible substances for further methane production. According to Jojoa-Unigarro and González-Martínez (2021), ethanol and acetic acid are the preferred substrates for methanization because of the potential inhibition caused by hydrogen partial pressure and acetogenesis of volatile fatty acids can be avoided. Although lactic acid needs to undergo acetogenesis before methanization, the hydrogen partial pressure does not affect this step. The novelty of this research lies in the use of UASB sludge and commercial yeast as inoculum to produce mainly ethanol and lactic acid through fermentation. This proposal also considers that undesired metabolites such as propionic and butyric acids are produced in lower concentrations when using these inocula for fermentation before methanization. The inevitable side production of acetic acid is welcome.
The methanization of the organic fraction of municipal solid wastes (OFMSW) consists of a complex series of biochemical reactions where different groups of microorganisms take an active part: The first step is the hydrolysis and acidification of complex organic molecules, and the second step is the methanization of the previously through fermentation produced metabolites. Most of the products from these fermentation processes are short-chain organic acids, alcohols, hydrogen, and carbon dioxide, required for the subsequent methanization. The fermentation depends on several variables: pH, temperature, inoculum composition, and products concentration [1]. Lin et al. [2] report that controlling the initial pH is essential to direct the fermentation to produce specific metabolites.
One limitation of the anaerobic digestion of OFMSW is that readily biodegradable substances can lead to acidification, decreasing pH, and inhibition of the microorganisms [3]. The main advantages of the two-stage systems are that acidification can be controlled in the first stage, allowing better control in the second stage, the methanization [4, 5, 6]. pH plays a vital role in the selectivity of the metabolites produced during fermentation. Under slightly neutral pH values, the fermentation predominates to propionic and butyric acids [7]; ethanolic fermentations are favored at pH between 5 and 5.4. Some other authors report that the best pH for ethanolic fermentation is near 4 where acid production decreases drastically [8, 9, 10, 11]. The solids retention time (SRT, for continuous and semi-continuous systems) or reaction time (for batch processes) is also another critical factor affecting ethanol production; adequate SRT or reaction time is relatively short, between 12 and 24 hours [10, 11, 12].
Zheng et al. [1] propose four different types of acidogenic fermentation: Ethanolic, butyric, mixed acids, and propionic. The ethanolic fermentation was defined when the proportion of ethanol (as COD) is higher than 30%, and the sum of acetate and ethanol is higher than 50%. In acidogenic reactors, the production of volatile fatty acids (VFA) has been the center of many research projects; few articles deal with the production of alcohols and lactic acid when fermenting OFMSW [13, 14, 15]. Ren et al. [16] pointed out that alcohol production is an exciting topic, but he concluded that during the production of hydrogen from OFMSW. Ethanol was not included in ADM1 because it has limited influence in one-step, low organic rate systems [17]. Nevertheless, ethanol is common in systems operated under high organic loading rates [1]. Working with batch reactors fed with food wastes, Jiang et al. [7] found that, at mesophilic temperature, when the pH was not controlled, the ethanol concentration was 11.75 gCOD/L, representing 66.6% of all the metabolites from the fermentation.
Pipyn and Verstraete [18] observed that ethanol and lactic acid formation during fermentation could improve subsequent methanogenesis because these intermediaries provide more energy to methanogens than other substances. Therefore, in a two-stage anaerobic digestion system, ethanol and lactic acid production needs to be favored to improve methane production. This could reduce the problems of acid inhibition as VFA production would decrease. Pipyn and Verstraete [18] report the Free Energy (∆G0) per mol CH4 recovered by archaea during methane production from different fermentation metabolites: Acetic acid, -31.0 kJ/mol, propionic acid, -32.3, butyric acid, -32.7, ethanol, -59.5, and lactic acid, -68.8. The energy recovered from the methanisation of ethanol and lactic acid double the one from acetic, propionic, and butyric acids.
Zheng et al. [1] propose that a controlled fermentation of OFMSW to produce preferably lactic acid and ethanol can benefit by reducing the risks of acidification during subsequent methanization. They also consider that ethanolic fermentation can stabilize a methanogenic reactor as an essential objective. Using fermented fruits and food wastes, with pH near 4, and ethanol concentrations of 12.5 gCOD/L, Shen et al. [21] report better and stable operation than when the methanogenic reactor was fed with an acetic-acid-rich substrate.
Considering that lactic acid and ethanol are thermodynamically desirable for later methane production [7, 19, 20], the main objective of this work was to favor the fermentation of OFMSW for better production of lactic acid and ethanol. Three different inocula were used: Naturally occurring microorganisms (NOM), yeast (Saccharomyces cerevisiae), and UASB sludge from a large brewery in Mexico City. Further analysis and methanization tests were performed to determine the effectivity of the fermentation as the first step of anaerobic digestion.
Reactor for Semi-continuous Fermentation
A 3-liter reactor with an active volume of 1.5 L was operated at 35°C under anaerobic conditions. The reactor has a twin-bladed mechanical mixing (Bioprocess Control, Sweden) and automatic pH control consisting of an electrode, a control device, and two peristaltic pumps for solutions of NaOH 2 M and phosphoric acid 2 M. pH was adjusted continuously between 4.0 and 4.1.
Daily, the fresh substrate was prepared with OFMSW suspended in tap water for a concentration of 40 gVS/L. One hundred mL were used for characterization, and the rest to feed the reactor. From the 1.5 L effective reactor volume, 1.35 L were extracted daily and replaced with the freshly prepared OFMSW suspension. Only 150 mL of the reactor content was left as inoculum for the next batch. Wu et al. [5], Zhou et al. [6], Shen et al. [21], and Zhao et al. [11] worked with substrate concentrations between 4 and 8% solids to avoid product inhibition. They also adjusted the reaction times between 12 and 36 hours as they observed the highest lactic acid and ethanol production. During feeding, the contents of the reactors were thoroughly mixed for 30 seconds to guarantee homogenization. Through OFMSW feeding, the solids retention time (SRT) was adjusted to 1.1 days.
The reactor operation consisted of three experiments, requiring six days every one of them: 1) OFMSW fermentation without inoculation (naturally occurring microorganisms, NOM); 2) inoculation with Saccharomyces cerevisiae, and 3) inoculation with anaerobic granular sludge from a UASB wastewater treatment facility of a large brewery in Mexico City. For the inoculation in experiments 2 and 3, 10 gVS of every inoculum was used [10].
Analytical Determinations
Total and dissolved COD, total and volatile solids, and pH were determined according to Standard Methods [22]. Methanol, ethanol, and VFA (acetic, propionic, isobutyric, butyric, isovaleric, valeric, hexanoic, and heptanoic acids) were determined using an Agilent gas chromatograph, model 8610, equipped with a flame ionization detector (FID), under the following conditions: one minute at 40°C with a ramp of 10°C per minute to reach 200°C. A Stabilwax column - DA was used with hydrogen as a carrier. Samples were previously filtered using 0.22 µm cellulose filters. Lactic acid was determined using the spectrophotometric method proposed by Borshchevskaya et al. [23].
Organic Fraction of Municipal Solid Waste (OFMSW)
The source-separated OFMSW was collected at the Coyoacán transfer station in Mexico City, where sampling was made according to the quartering method [24]. Approximately 100 kg from every one of 11 trucks were separated, and about one ton was thoroughly mixed using a skid-steer loader and shovels. After that, the quartering method was applied two times to reduce the amount to approximately 200 kg. Undesired materials, such as plastic bags, stones, and wood, were hand separated; the remaining "clean" OFMSW was distributed in two-liter freezing bags and frozen at -20 °C. From this amount, 20 kg were overnight defrosted at 4°C and, using a grain grinder, particles under 5 mm were obtained. OFMSW was characterized for total solids (TS), 279±8 g/kg, volatile solids (VS), 211±4 g/kg, chemical oxygen demand (COD), 389±48 g/kg, and total Kjeldahl nitrogen (TKN), 5.1±0.4 g/kg. All determinations were made according to Standard Methods [22].
Inocula
The anaerobic granular sludge was collected in a UASB wastewater treatment facility of a large brewery in Mexico City. To separate the soluble substances ("sludge washing") from the granular sludge, 400 ml were centrifuged for 7 min at 3,600 RPM. The supernatant was discarded and replaced with tap water to complete the 400 mL; after thorough mixing, the suspension was again centrifuged. This procedure was repeated three times.
The yeast was commercial lyophilized baking 100% Saccharomyces cerevisiae. The characteristics were determined to be 954±5 2 gTS/kg and 731±8 gVS/kg.
Methanization
The last samples of the 6-day experiments were placed in an automatic biogas production monitoring system (AMPTS II, Bioprocess Control, Sweden) to evaluate the effects of fermentation on methane production. For the methanization, granular sludge from a large UASB wastewater facility from a large brewery was used as methanogenic inoculum. An 8 gVS inoculum was placed in every 500 mL flask. Using the digestates from the fermentation experiments as substrate, four concentrations were adjusted at 1.25, 2.5, 5.0, and 10.0 gVS/L. After placing the inoculum and substrate in every flask, to control the pH at 7.0, a 0.1 M phosphate buffer was used as dilution water in every flask to reach a final volume of 400 mL.
At the end of the methanization test, samples were taken and analyzed for alcohols and VFA, including lactic acid.
The biogas composition (CO2 and CH4) was determined using a gas chromatograph (SRI 8610c) equipped with a thermal conductivity detector, stainless steel column packed with silica gel (8600-PK1A), helium as carrier gas with a flow rate of 27 mL/min. The detector temperature was 150 °C.
Modeling
A modified Gompertz model, non-linear, Eq. 1 [25], was used to corroborate the experimental methanization values such as the maximal production rate, methane yield, specific methane production, and latency time (lag phase). The model corresponds to a descriptive sigmoid function.

Y(t) is the accumulated methane specific production (mL/gVS), t is time (days), Pmax the maximum methane production potential (mL/gVS), Rmax the maximum methane production rate (mL/gVS·d), λ is the lag phase (days), and ℯ is the Euler mathematical constant, 2.7183. The model adjustment was made using the Statistica 7.0 software, minimizing the error between calculated and experimental values. The adjustment level was the quadratic correlation coefficient (R2).
Fermentation with Different Inocula
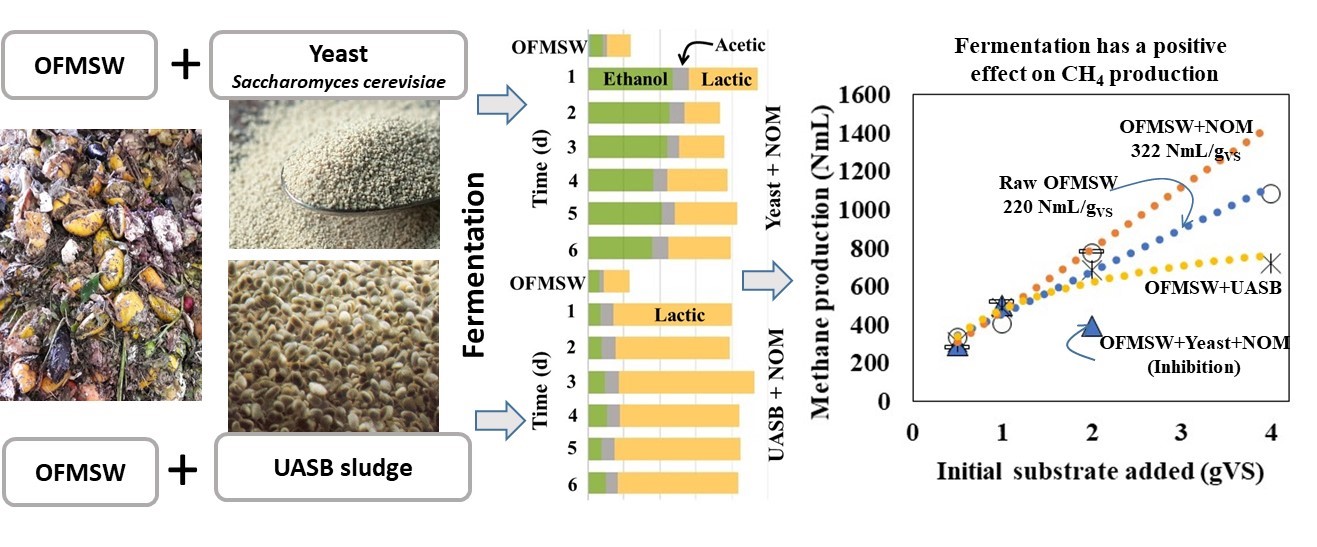
Figure 1 shows the different metabolites produced during the OFMSW fermentation with different inocula. The semi-continuous operation of the reactors resulted in an SRT of 1.1 d. The test took six days for every inoculum, and samples for lactic acid, VFA, and alcohols were taken every day. As the fresh OFMSW was collected at the origin and transported to the transfer station, several hours passed, incipient fermentation was expected during this time; these are the values indicated as OFMSW at day zero with 6.9±0.7 gCOD/L where lactic acid and ethanol are the main components. Small concentrations of acetic acid were detected in fresh OFMSW.
Naturally Occurring Microorganisms (NOM). Ethanol reached a maximal concentration of 5.3 gCOD/L on the second day, and then it slowly began to decrease to 4.5 gCOD/L on day 6. As the ethanol concentration decreased, acetic acid concentration increased with time to 4.1 gCOD/L on day 6. Lactic acid reached 15.5 gCOD/L on day 4, and it did not change during the next two days. CODof OFMSW at day zero was 6.9 g/L, and it increased to 20 g/L on day 4, indicating that dissolved substances contribute to the formation of acids and alcohols.
--- Figure 1 ---
Yeast. The highest ethanol concentration was reached on the first day with 12 gCOD/L, and then it decreased to 8.5 gCOD/L on the sixth day. Lactic acid reached a maximal concentration on the first day with 9.8 gCOD/L, dropped significantly on the second day, and then it increased steadily until day six; the final value was 9.5 gCOD/L. The acetic acid concentration did not vary significantly over the six days, maintaining values between 2.0 and 2.5 gCOD/L.
UASB Sludge. Ethanol maintained values between 1.6 and 2.3 gCOD/L from days one to six. The same behavior and values were observed for acetic acid. The surprise was the lactic acid production with a maximum of 19.6 gCOD/L on the third day. Lactic acid concentrations do not show a tendency as the concentration remains without significant changes over the experimental time.
The most important products from the fermentations were ethanol and lactic acid, corresponding to almost 90% of all products; the other 10% was acetic acid. For lactic acid, the highest production was 80% of the total metabolites, corresponding to the fermentation with UASB sludge, then with the native occurring microorganisms, 63%, and finally, with yeast, 41% was detected. The highest ethanol production was obtained with yeast as inoculum with 50% of the total produced metabolites, then with NOM with 28%, and finally with UASB sludge with 12%. Like the results of Zheng et al. [1], this research observed that, during the fermentation with yeast, ethanol concentration slightly decreased as lactic acid concentration increased. In all three cases, acetic acid represented between 8 and 10% of all metabolites without identifying a tendency in the production.
The microorganisms were seeded only once at the beginning and, after every feeding procedure, approximately 10% of the original seed remained in the reactor for the next batch. Figure 1 shows that for yeast+NOM, the ethanol production decreases and lactic acid increases with time: On one side, these results are associated with the wash-out of the microorganisms with every feeding procedure and, on the other, with increasing the NOM, slowly returning the NOM original values. This behavior was not observed for UASB sludge+NOM; the concentrations of ethanol, acetic acid, and lactic acid did not report significant changes during the six days fermentation. For only NOM, the activity of the lactate fermenting microorganism increased with time, and the ethanol and lactic acid concentration did not change significantly over time.
Independently of the inoculum and operation conditions (SRT, 1.1 d and constant pH at 4.1), acetic acid was not the preferred fermentation. For all three cases, the generation was approximately the same after six days. Jankowska et al. [26] evaluated a reactor under HRT of 5, 10, and 15 d and pH from 4 to 12, and they concluded that, under pH 4 and 5, and HRT of 5, the VFA production decreases. Wu et al. [10] worked a reactor under pH of 3 and 4, and they managed selectivity for ethanol and very low for acetic acid. Cheah et al. [14] analyzed the fermentation at a pH of 4 and obtained VFA where acetic acid represented 67% of all VFA; the difference was butyric and caproic acids. This research's average acetic acid production was 2.15 gCOD/L, similar to the reported in the previously mentioned articles.
Fermentation Rates
The fermentation rate can be calculated as a CODfraction of the maximum CODvalue. Figure 1 presents the fermentation rate for total metabolites and individually.
Naturally Occurring Microorganisms (NOM).For NOM, the total fermentation rate is low during the first two days, and then it increases drastically to almost 100%; after that, the rate decreases slowly to 70% at the end of the test. Lactic acid rapidly increased on the third day, and it decreased until the end of the test. The fermentation rate of ethanol increased the first day rapidly to 34%, remained unchanged on the second day, and then steadily decreased to reach a final value of 15%. Contrary to the other metabolites, acetic acid decreased during the first two days, increased to 10%, and remained at 9% the last three days. Although 90% of the reaction volume was replaced every day, the microorganisms increased their activity during the test. Lactic acid was the preferred fermentation.
Yeast + NOM.The total fermentation rate oscillated between 60 and 80%. Ethanol presented the highest fermentation rate on the first day with 37%, and then it decreased slowly to 32% on the last day. Lactic acid increased on the first day to decrease on the second, and then it increased again on the fourth day to slightly decrease from 34 to 21%. Acetic acid remained with almost no changes during the whole experiment, with an average of 7%. These results show that the yeast did not grow sufficiently to increase its concentration and that it was slowly washed from the reactor. In the beginning, there was a maximal yeast concentration, and ethanol increased rapidly, but, with time, the concentration and fermentation rates decreased as the yeast was washed out of the system.
UASB Sludge+NOM.The fermentation rate with UASB sludge is the highest of the three. The total fermentation rate increased the first day rapidly to 81%; on day three, it reached 100%, and then it decreased slowly to 93% at the end of the test. With lower values, 78% as maximum, lactic acid fermentation behaved similarly, indicating that both lactic acid concentration and fermentation rate were highest when fermenting with UASB sludge. The fermentation rates of acetic acid and ethanol remained low, with average values near 10% during the six days of the test. These conditions promote lactic acid fermentation.
Soomro et al. [27] investigated the OFMSW fermentation without pH control and, unfortunately, did not analyze for ethanol and lactic acid as they discarded their presence as a possibility. They conclude that at pH values of 4, the fermentation stopped and, after three days, the butyric fermentation took place. Gameiro et al. [15] studied OFMSW fermentations without inoculum under different pH values, and they reported that acid fermentation stopped at a pH of 4 with a total fermentation rate of 15%. After studying the fermentation of food wastes (vegetables and fruits) under pH values of 4, 5, and 6, at 37°C, Zheng et al. [1] conclude that at pH of 4, the fermentation rate for ethanol was 97.5% and the rest was acetic acid. Wu et al. [10] worked with a CSTR at 35°C, HRT 3 d, and pH from 4 to 7, using food waste as substrate; they report that at pH 4 the fermentation was 88% to ethanol and the rest acetic acid.
The previously reported authors' results differ from those in this research because some authors did not analyze lactic acid and/or ethanol, and this discussion is only partial. In this research, lactic acid was predominant when NOM and UASB sludge were used as inoculum, and ethanol was the primary metabolite when yeast was added as seed at the beginning.
According to the specialized literature, there are two metabolic routes for lactic acid production: Homofermentative and heterofermentative (Eq. 2 and 3) [10]. The products from the homolactic fermentation are mainly lactic acid, and the ones from the heterolactic fermentation are equimolar quantities of CO2, lactate, and acetate or ethanol [28]. The average concentrations of acetic acid, ethanol, and acetic acid in the case of the fermentation with NOM were 30±5, 51±6, and 132±4 mM, respectively; in the case of the fermentation with yeast+NOM, the values were 32±4, 108±10, and 98±15 mM, respectively; the last case, the fermentation with UASB sludge+NOM reported 28±2, 22±3, and 178±11 mM, respectively. Table 1 shows the final concentrations of ethanol and lactic and acetic acids after the fermentation for six days. The ratio of lactic acid to ethanol or acetic acid during the fermentation with NOM was, for all conditions, higher than one, proving a homolactic fermentation because the production of acetic acid or ethanol is not related to the production of lactic acid. During the fermentation with yeast+NOM, the molar concentration of ethanol and lactic acid are similar, and the lactic acid to ethanol ratio is 0.91 indicating heterofermentative, but Saccharomyces cerevisiae ferment only to ethanol. There must have been simultaneous participation of heterofermenting and homofermenting microorganisms present as equimolar quantities of ethanol and lactic acid were produced (Table 1).

--- Table 1 ---
Table 2 shows total and soluble COD at the end of the 6-day fermentation tests for the three cases. For all three cases, total COD decreased during the six days from 9% in the fermentation with UASB sludge to 14% for the fermentation with NOM. Simultaneously, soluble COD increased in all three cases: 16% for the fermentation with yeast+NOM, 9% for NOM, and 5% for UASB sludge+NOM.
Also, from Table 2, the TS decreased 21% for the fermentation with NOM, and 4% for both fermentations with yeast+NOM and UASB sludge+NOM. For the fermentation with NOM, the VS decreased 25%, 19% for yeast+NOM, and 10% for UASB sludge+NOM. The solids decrease indicates lower hydrolysis rates for all three cases. Kandylis et al. [29] report deficient hydrolytic processes when the pH is low, especially under 5.5, where the enzymatic activity is also very low and inhibits microbial metabolism. Gameiro et al. [15] confirm that fermentation at pH of 4 deactivates the hydrolytic processes as it inhibits the microbial processes.
--- Table 2 ---
Methanization of Digestates from the Previous Fermentation
After six days of fermentation with different inocula, the resulting digestates were analyzed for methane production at various substrate concentrations (Figure 2). Raw OFMSW was used for comparison, and the test ran for 30 days. As a general observation, methane production increased with increasing substrate concentration, but inhibition was observed at higher substrate concentrations.
--- Figure 2 ---
Methane Production from Raw, Unfermented OFMSW (reference substrate)
This test was conducted to reference how much methane can be obtained directly from the methanization of the unfermented OFMSW. Figure 2 shows the methane production curves during 30 days. The lowest curve is the blank, only inoculum, and the other curves indicate increasing methane production with increasing substrate initial concentration. During the first days, methane production is fast, and then the production rate decreases showing a second stage, typical for acetogenesis [30]. Figure 3 indicates that the final methane production is directly proportional to the initial substrate concentration with a good correlation coefficient.
Methane Production from OFMSW Fermented only with Naturally Occurring Microorganisms (NOM)
As in the previous case, methane production started fast, and then it decreased to show a prolonged lag-phase (diauxic behavior) for the two highest concentrations. The highest substrate concentration presents a parallel production to the blank and, after approximately ten days, it increased significantly to finish the production on day 27. The final values after 30 days are similar to the ones of the raw unfermented OFMSW. Figure 3 shows a linear behavior between final methane production and initial substrate concentration. These last values are almost the same as those of the final methane production of raw unfermented OFMSW.
--- Figure 3 ---
Methane Production from OFMSW Fermented with Yeast and NOM
The two lower initial substrate concentrations (1.25 and 2.5 gVS/L) were similar to the two previously discussed cases. The two higher concentrations (5 and 10 gVS/L) had different patterns, indicating substrate inhibition. The 5 gVS/L case presented similar values as the case with 2.5 gVS/L and final value, considering that the 30-day test was not enough for the reaction. For the 10 gVS/L case, after three days, the reaction stopped and, after ten more days, the methane production suddenly reactivated; unfortunately, a problem with the reaction cell prevented methane monitoring after day 22. Figure 2 shows that the two lower initial substrate concentrations correspond with the methane production of the unfermented OFMSW and the OFMSW fermented only with NOM. The value for 5 gVS/L is low, indicating inhibition, and the value for 10 gVS/L is missing.
Methane Production from OFMSW Fermented with UASB Sludge and NOM
As in the previous cases, the methane production during the first two days was fast, and then it decreased considerably to rates similar to the blank during several days. Only the methane production of the highest initial substrate concentration stopped after three days. Then, it began suddenly on day 12 and stopped short on day 17; it increased again to stop on day 26. The two lowest substrate concentrations continued producing methane at the same rate as the blank after day 12. The final values are shown in Figure 3.
During the first seven days, all the curves in Figure 2 show similar behavior. The digestate from the fermentation with yeast+NOM presents the lowest methane production during the first seven days. The lowest substrate concentration reports the lowest methane production rates, followed by the next higher substrate concentration. As the substrate concentration increases, methane production varies according to the substrate type. The methane production curves show two stages, and the second corresponds to acetogenesis followed by methanogenesis [30].
An evident inhibition is observed for the 10 gVS/L concentration as methane production stops after three days for the digestates from the yeast+NOM and UASB sludge+NOM fermentations. In the case of the yeast+NOM, the methane production reactivates drastically after 17 days, and, unfortunately, a failure in the methane counter cell did not allow to follow the further development of the curve. It took 11 days for the UASB+NOM to reactivate with a sudden increase in methane production; the curve indicates instability till the end of the experiment.
Figure 3 shows the values for the final methane production, calculated from Figure 2. The final methane produced is plotted versus the substrate added at the beginning of the test. The best fit for a straight line is for unfermented OFMSW, meaning that the methane production from different initial substrate concentrations is directly proportional to the substrate concentration, and no inhibition could be detected. At lower substrate concentrations, the methane production from the digestates fermented with NOM, yeast+NOM, and UASB sludge+NOM are higher than the unfermented OFMSW.
The specific methane production, obtained through Figure 3, considers only the lower three substrate concentrations. As seen in Figures 2 and 3, the final values for the 4 gVS tests are, except for the unfermented OFMSW, lower than expected. Inhibition is the cause of this behavior. In the case of the fermentation with yeast+NOM, only the two lowest values are considered to behave consistently with the other cases. The values that do not fit in the lines are lower than expected, consistent with inhibition. In subsequent sections of this article, it will be shown that this inhibition is caused by propionic acid.
Table 3. shows the specific methane production calculated from Figure 3. Theoretically, the intercept corresponds to the blank methane production. Jojoa-Unigarro and González-Martínez [30] demonstrated that endogenous methane production strongly depends on the substrate, leading to a wrong interpretation when considering endogenous production as a constant. Analyzing the scale in the y-axis and intercepts in Figure 3, and considering the analytical error, the calculated intercepts of unfermented OFMSW and fermented with UASB sludge+NOM are close. The digestate from the fermented OFMSW with NOM is higher (322 NmL/gVS) than the other two, indicating higher endogenous methane production.
--- Table 3 ---
Discussion on Methane Production from Fermented Organic Solid Wastes
Table 4 compares the results obtained in this research with other authors. In most cases, the methane productions are similar among the authors, and, in a few instances, the production is higher than most. The primary discrepancies in the values reported in Table 4 are based on the origin and composition of the wastes. Working with OFMSW from the same source as this research, Figueroa-Escamilla and González-Martínez [31] report higher amounts of lignocellulosic compounds from garden wastes included in the OFMSW. The authors cited in Table 4 worked mainly with fruits and vegetable wastes containing more elevated amounts of sugars and other readily biodegradable substances. Unlike this research where different variables were tested, several authors determined the methane yield based on limited operational conditions; in some cases, they report the values from liquid wastes where the hydraulic retention times (HRT) were relatively low [3, 32, 33]. These last authors report methane production from 300 to 350 NL/kgVS, which is higher than this research (max. 196 NL/kgVS). Liquid wastes are known to contain higher concentrations of readily biodegradable substances as most of the complex substances remain in the solid fraction.
--- Table 4 ---
Working with CSTR and relatively low organic loading rates and in one-stage and two-stage anaerobic digestion, Baldi et al. [34] obtained the highest methane production rates with 694 and 705 NL/kgVS, respectively, where the highest value corresponds with the two-stage anaerobic digestion. These values are much higher than those reported by Campuzano and González-Martínez [35] and those obtained in this research.
Baldi et al. [34], Ma et al. [36], and Valentino et al. [37] evaluated the anaerobic digestion process in two stages and compared their results with the methane production in one stage using food wastes as substrate. It is important to note that the reported results in Table 4 contain similar process variables to those in this research. For example, all values were obtained under mesophilic temperature, and SRT was from 15 to 30 days. Some authors report results from batch experiments and others from CSTR. Comparing the results of the one-stage process, in the two-stage process, they managed an increase in methane production between 0.5 and 10%. This research achieved a 5% increase when the substrate concentrations were relatively low because the inhibition caused by higher substrate concentrations reduced methane production.
Using maize, fruits, and vegetable wastes in a two-step process, Schievano et al. [38] noted that the advantages of the two-stage process result in a better energy recovery; they report an increase of 23 to 43% in energy recovery compared to the one-step process only when operating under lower organic loading rates. Under pH of 6.6, Valentino et al. [37] conclude that under these conditions, propionic and valeric acid fermentations are predominant, representing both acids 50% of the total metabolites. They also conclude that methanogenesis after the fermentation must be run under lower organic loading rates to allow the uptake of propionic and valeric acids through acetogenesis.
Yu et al. [39] report similar methane productions to this research (184 vs. 196 NL/kgVS). The main differences are that Yu et al. [39] worked at 37°C instead of 35°C, and the reaction time was 20 days instead of 30 days. The general conclusion is that a two-stage process is more flexible than a one-stage process. Higher methane productions are achieved with continuous reactors, followed by semi-continuous and, finally, with discontinuous systems. To achieve higher methane production, methanization needs to be run under lower organic loading rates independently of the inoculum used for fermentation.
Modeling with the Modified Gompertz Equation
For the Gompertz modeling, the methane produced was transformed in specific methane production by dividing the produced methane by the substrate concentration (Figure 1). Using the program Statistica 7.0 a non-linear fit was plotted (Figure 4) to minimize the differences between calculated and experimental values. As a result, the parameters Pmax, Rmax, and λ can be calculated (Eq. 1).
--- Figure 4 ---
According to the Gompertz modeling, Table 5 shows the specific methane production according to the initial substrate concentration of the different fermented digestates and raw, unfermented OFMSW. Pmax is the maximal methane yield or specific methane production; Rmax is the maximum methane production rate, and λ is the latency time. The specific methane production is inversely proportional to the substrate concentration for all tested substrates, the experimental data, and the calculated values. For every one of the tested digestates, the highest methane production corresponds to the concentration of 1.25 gVS/L. The most increased methane production corresponds to the fermentation with yeast+NOM with 707 NL/kgVS, and it represents only 6% more than the methane production of unfermented OFMSW.
The lowest specific methane production corresponds to unfermented OFMSW at 1.25 gVS/L with 662 NL/kgVS; with increasing substrate concentration to 2.5 gVS/L, the methane production significantly decreased to 421 NL/kgVS. At a substrate concentration of 10 gVS/L the specific methane production fell to 269 NL/kgVS.
At a substrate concentration of 1.25 gVS/L, the digestate from the fermentation with UASB sludge+NOM, produced 671 NL/kg at 10 gVS/L, and this production decreases as the substrate concentration increases. For the digestate from yeast+NOM, the specific methane production decreases at a substrate concentration of 2.5 gVS/L, and it decreases further to 157 NL/kgVS as the substrate concentration increases to 10 gVS/L.
Using the modified Gompertz model, Figure 4 compares the calculated with the experimental values. The correlation coefficients are between 0.98 and 0.99 (Table 5). Comparing Pmax with the observed values, the model overestimates between 0.5 to 14%, the methane production cases, and substrate concentrations. For the methane production from the USAB sludge+NOM digestate, the model overestimates 37 to 49% of the two highest substrate concentrations values. Similar is the behavior for unfermented OFMSW at the highest concentration, where the overestimation is 41%. These results indicate that the adjustments made by the modified Gompertz model fit better for the lower substrate concentrations, independently of the case.
The Gompertz model is a sigmoid function, and it fits better when anaerobic digestion consists of only one step when the methane production is used as a variable. The model indicates that the curve would stabilize when reaching the methane production of 384 NL/kgVS (Table 5). At the higher concentrations of 5 and 10 gVS/L, Figure 4 shows that the fitted curve does not pass near all the experimental data, and it tends to show the data as a straight line.
Pmax indicates that the methane production rate decreases with increasing substrate concentration (Table 5). The curves in Figure 5 shows that the methane production during the first days is faster as the substrate concentration decreases. Only for the digestate from the fermentation with yeast+NOM, the behavior cannot be predicted with the experimental values, but the Gompertz adjusted curve allows to predict the behavior. It can be concluded that the three cases proposed in this research produce methane faster than the unfermented OFMSW at lower substrate concentrations.
The latency time is essential for the evaluation of the anaerobic digestion process. It reflects the time the microorganisms require to adapt to the new conditions, including the new substrate. Table 5 shows that, except for two experiments with a substrate concentration of 10 gVS/L, all λ values have a negative value, indicating that the microorganisms began methane production immediately after being put in contact with the substrate. The positive values in the cases of the fermentation with NOM and yeast+NOM under 10 gVS/L substrate concentration suggest a substrate inhibition. According to Chu and Barnes [40], the negative values correspond to a well-adapted inoculum and a readily biodegradable substrate.
Metabolites Remaining after Methanisation
After the methanisation of the different fermented digestates, analysis for other possible metabolites was made after the 30 days of the test. Figure 5 shows the relative concentrations, as COD, versus the initial substrate concentrations, as VS. Identified were methanol, ethanol, and acetic, propionic, butyric, and isobutyric acids. Surprisingly, several of these metabolites remained after 30 days of methanization.
--- Figure 5 --
Unfermented OFMSW. Unfermented OFMSW was subjected to methanization to have a reference. The acetic acid production increased inversely with the substrate concentration, and the production of propionic and butyric acids increased with substrate concentration. Ethanol remained relatively low independently of the initial substrate concentration.
Digestate from the Fermentation with NOM. The digestate from the fermentation with NOM produced the highest concentrations of acetic acid. Acetic acid production was inversely proportional to the initial substrate concentration, and propionic and butyric acids increased substantially with increasing substrate concentration.
Digestate from the fermentation with Yeast+NOM. The highest concentration of propionic acid remained after the methanization of the digestate from the fermentation with yeast; the propionic acid production increased substantially with the substrate concentration. With increasing propionic acid, butyric acid also increased but with lesser concentrations. Ethanol and acetic acid remain in lower concentrations, although a total consumption could have been expected after 30 days of methanization.
At the end of the methanization, the digestate from yeast+NOM shows lower concentrations of ethanol and acetic and butyric acids at lower substrate concentrations (Figure 5); at higher substrate concentrations, at the end of the methanization, propionic acid is also present with 1.1 gCOD/L. This explains why the methane production decreased under the higher substrate concentrations: Propionic acid is an inhibitor of methanization. Jojoa-Unigarro and González-Martínez [30], in their research about acetogenesis, report that propionic acid concentrations above 300 mgCOD/L partially inhibit methanization and that, concentrations above 1,200 mgCOD/L the methane production is inhibited under the values of the blank. When lactic acid is present, acetogenesis produces propionic acid, and methanization can be inhibited. In their study about anaerobic digestion in two stages, Shen et al. [21] report that when the organic loading rate is not adjusted correctly, higher concentrations of metabolites will be produced, and VFA and alcohols will cause inhibition during methanization. Significantly, propionic and butyric acids need to be considered to reduce the adverse effects of inhibition on the methanization process.
Digestate from the Fermentation with UASB Sludge+NOM. Figure 5 shows that, at the end of the methanization, the concentration of propionic acid increased drastically with substrate concentration. The methanization from this digestate shows inhibition caused by propionic acid at the highest concentration of 10 gVS/L. Ethanol was present at lower substrate concentrations and butyric acid only at the highest concentration. The combination of propionic and butyric acids at the higher concentration is the cause for the significant inhibition (Figure 3). The acetic acid concentration did not change significantly with substrate concentration, and isobutyric acid was detected only at the lowest substrate concentration.
-
OFMSW fermentation with naturally occurring microorganisms (NOM) and UASB sludge + NOM produce mainly lactic acid, 60% and 80% of total COD, respectively. Yeast + NOM fermented primarily in ethanol with 50%.
-
The highest hydrolysis rates were achieved with yeast + NOM with 66%. Total solids decreased 10% in six days, and the dissolved solids increased in the same proportion.
-
The methane production from the fermentation digestates increases with decreasing substrate concentration.
-
The highest methane production was obtained with the digestate from the OFMSW fermented only with NOM, followed by the digestate fermented with UASB sludge + NOM. The lowest specific methane production is the one with raw, unfermented OFMSW. Independently of the inocula, all the fermented digestates produced more methane than the unfermented OFMSW.
-
Methane production from the digestates from yeast + NOM increased with the initial concentration; propionic and butyric acids concentrations significantly increased with substrate concentration.
-
Propionic and butyric acids are the primary metabolites remaining after the methanization of the fermented digestates. After 30 days of methanization of the digestate from yeast + NOM, with increasing initial digestate concentration, propionic and butyric acids increased from 350 to 1,250 mgCOD/L; this indicates inhibition during methane production at higher substrate concentrations.
-
Modeling with the Gompertz equation correlates with the experimental data and indicates that inhibition increased with increasing substrate concentration, independently of the inocula used for previous fermentation.
-
The digestate from UASB sludge + NOM showed lower propionic and butyric acid concentrations than the other digestates; inhibition at higher substrate concentrations was detected.
-
The methane produced from raw, unfermented OFMSW was not inhibited; the propionic and butyric acids concentrations after the 30-day methanization were the lowest of all four cases.
Acknowledgment
This project was supported by the Mexican Centre for Innovation in Bioenergy-Cluster for Gaseous Biofuels (Centro Mexicano de Innovación en Bioenergía, Clúster en Biocombustibles Gaseosos) CONACYT project 247006, and the General Directorate for Academic Affairs (Dirección General de Asuntos del Personal Académico, DGAPA) of the National University of Mexico (UNAM), project IT101320. This research was performed at the Environmental Engineering Laboratory (LIA), Institute of Engineering, National University of Mexico. The graduate scholarship provided by CONACYT is acknowledged. The authors recognize the unconditional support in the laboratory and guidance through the theoretical supervision of Dr. Óscar González-Barceló.
CRediT authorship contribution statement
Investigation, conceptualization, and writing, G.D. Jojoa-Unigarro. Conceptualization, writing, and editing: S. González-Martínez
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Data availability statement in the manuscript
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
- Zheng, J., Tashiro, T., Wang, Q., Sonomoto, K.: Recent advances to improve fermentative butanol production: Genetic engineering and fermentation technology. J. Biosci. Bioeng. 119, 1–9 (2015). https://doi.org/10.1016/j.jbiosc.2014.05.023
- Lin, L., Wan, C., Liu, X., Lee, D.J., Lei, Z., Zhang, Y., Tay, H.: Effect of initial pH on mesophilic hydrolysis and acidification of swine manure. Bioresour Technol. 136, 302–308 (2013). https://doi.org/10.1016/j.biortech.2013.02.106
- Ganesh, R., Torrijos, M., Sousbie, P., Lugardon, A., Steyer, J.P., Delgenes, J.P.: Single-phase and two-phase anaerobic digestion of fruit and vegetable waste: Comparison of start-up, reactor stability and process performance. Waste Manag. 34, 875–885 (2014). https://doi.org/10.1016/j.wasman.2014.02.023
- Bouallagui, H., Lahdheb, H., Romdan, E., Ben, Rachdi, B., Hamdi, M.: Improvement of fruit and vegetable waste anaerobic digestion performance and stability with co-substrates addition. J. Environ. Manage. 90, 1844–1849 (2008). https://doi.org/10.1016/j.jenvman.2008.12.002
- Wu, C., Wang, Q., Yu, M., Zhang, X., Song, N., Chang, Q., Gao, M., Sonomoto, K.: Effect of ethanol pre-fermentation and inoculum-to-substrate ratio on methane yield from food waste and distillers' grains. Appl. Energy. 155, 846–853 (2015). https://doi.org/10.1016/j.apenergy.2015.04.081
- Zhou, M., Yan, B., Wong, J.W.C., Zhang, Y.,. . ., Part, A.: Enhanced volatile fatty acids production from anaerobic fermentation of food waste: A mini-review focusing on acidogenic metabolic pathways. Bioresour Technol. 248, 68–78 (2018). https://doi.org/10.1016/j.biortech.2017.06.121
- Jiang, J., Zhang, Y., Li, K., Wang, Q., Gong, C., Li, M.: Volatile fatty acids production from food waste: Effects of pH, temperature, and organic loading rate. Bioresour Technol. 143, 525–530 (2013). https://doi.org/10.1016/j.biortech.2013.06.025
- Taghizadeh-Alisaraei, A., Motevali, A., Ghobadian, B.: Ethanol production from date wastes: Adapted technologies, challenges, and global potential. Renew. Energy. 143, 1094–1110 (2019). https://doi.org/10.1016/j.renene.2019.05.048
- Wang, Y.F., Tan, L., Wang, T., Sun, Z.Y., Tang, Y.Q., Kida, K.: Production of ethanol from kitchen waste by using flocculating Saccharomyces cerevisiae KF-7. Environ. Technol. 38, 316–325 (2017). https://doi.org/10.1080/09593330.2016.1192224
- Wu, Y., Wang, C., Zheng, M., Zuo, J., Wu, J., Wang, K., Yang, B.: Effect of pH on ethanol-type acidogenic fermentation of fruit and vegetable waste. Waste Manag. 60, 158–163 (2017). https://doi.org/10.1016/j.wasman.2016.09.033
- Zhao, N., Yu, M., Wang, Q., Song, N., Che, S., Wu, C., Sun, X.: Effect of Ethanol and Lactic Acid Pre-fermentation on Putrefactive Bacteria Suppression, Hydrolysis, and Methanogenesis of Food Waste. Energy Fuels. 30, 2982–2989 (2016). https://doi.org/10.1021/acs.energyfuels.5b02779
- Lim, S.J., Kim, B.J., Jeong, C.M., Choi, J.dal-rae, Ahn, Y.H., Chang, H.N.: Anaerobic organic acid production of food waste in once-a-day feeding and drawing-off bioreactor. Bioresour Technol. 99, 7866–7874 (2008). https://doi.org/10.1016/j.biortech.2007.06.028
- Dionisi, D., Anderson, J.A., Aulenta, F., McCue, A., Paton, G.: The potential of microbial processes for lignocellulosic biomass conversion to ethanol: a review. J. Chem. Technol. 90, 366–383 (2015). https://doi.org/10.1002/jctb.4544
- Cheah, Y.K., Vidal-Antich, C., Dosta, J., Mata-Álvarez, J.: Volatile fatty acid production from mesophilic acidogenic fermentation of organic fraction of municipal solid waste and food waste under acidic and alkaline pH. Environ. Sci. Pollut Res. 26, 35509–35522 (2019). https://doi.org/10.1007/s11356-019-05394-6
- Gameiro, T., Lopes, M., Marinho, R., Vergine, P., Nadais, H., Capela, I.: 2016. Hydrolytic-Acidogenic Fermentation of Organic Solid Waste for Volatile Fatty Acids Production at Different Solids Concentrations and Alkalinity Addition. Water Air Soil Pollut. 227, 391 (2016). https://doi.org/10.1007/s11270-016-3086-6
- Ren, N., Wang, B., Huang, J.C.: Ethanol-type fermentation from carbohydrate in high rate acidogenic reactor. Biotechnol. Bioeng. 54, 428–433 (1997). https://doi.org/10.1002/(SICI)1097-0290(19970605)54:5<428::AID-BIT3>3.0. CO;2-G
- Batstone, D.J., Keller, J., Angelidaki, I., Kalyuzhnyi, S.V., Pavlostathis, S.G., Rozzi, A., Sanders, W.T.M., Siegrist, H., Vavilin, V.A.: The IWA Anaerobic Digestion Model No 1 (ADM1). Water Sci. Technol. 45, 65–73 (2002). https://doi.org/10.2166/wst.2002.0292
- Pipyn, P., Verstraete, W.: Lactate and ethanol as intermediates in two-phase anaerobic digestion. Biotechnol. Bioeng. 23, 1145–1154 (1981). https://doi.org/10.1002/bit.260230521
- Hiu, Z., Gao, M., Yu, M., Zhang, M., Zhang, Sh, Wu, Ch, Tashiro, Y., Wang, Q.: Methane production from food waste via mesophilic anaerobic digestion with ethanol pre-fermentation: Methanogenic pathway and microbial community analyses. Bioresour Technol. 297, 122450 (2020). https://doi.org/10.1016/j.biortech.2019.122450
- Zhang, B., Zhang, L.-L., Zhang, S.-C., Shi, H.-Z., Cai, W.-M.: The Influence of pH on Hydrolysis and Acidogenesis of Kitchen Wastes in Two-phase Anaerobic Digestion. Environ. Technol. 26, 329–340 (2005). https://doi.org/10.1080/09593332608618563
- Shen, F., Yuan, H., Pang, Y., Chen, S., Zhu, B., Zou, D., Liu, Y., Ma, J., Yu, L., Li, X.: Performances of anaerobic co-digestion of fruit & vegetable waste (FVW) and food waste (FW): Single-phase vs. two-phase. Bioresour Technol. 144, 80–85 (2013). https://doi.org/10.1016/j.biortech.2013.06.099
- Standard Methods for the Examination of Water and Wastewater. 23rd edition, American Public Health Association/American Water Works Association/Water Environment Federation, Washington DC, USA: (2017)
- Borshchevskaya, L.N., Gordeeva, T., Kalinina, A.N., Pavlovich, S.: Spectrophotometric determination of lactic acid. J. Anal. Chem. 71, 755–758 (2016). https://doi.org/10.1134/S1061934816080037
- ASTM Standard Test Method for Determination of the Composition of Unprocessed Municipal Solid Waste. 2016. ASTM D5231-92 ASTM International, West Conshohocken, PA
- Venkataramana, G., Yalini, A., Nagamany, N., Johnson, D.C.: Modeling dark fermentation for biohydrogen production: ADM1-based model vs. Gompertz model,International. J. Hydrogen Energy. 35, 2, 479–490 (2010). https://doi.org/10.1016/j.ijhydene.2009.11.007
- Jankowska, E., Chwiałkowska, J., Stodolny, M., Oleskowicz-Popiel, P.: Effect of pH and retention time on volatile fatty acids production during mixed culture fermentation. Bioresour Technol. 190, 274–280 (2015). https://doi.org/10.1016/j.biortech.2015.04.096
- Soomro, A.F., Abbasi, I.A., Ni, Z., Ying, L., Liu, J.: 2020. Influence of temperature on enhancement of volatile fatty acids fermentation from organic fraction of municipal solid waste: Synergism between food and paper components. Bioresource Technology, 304, 2020, 122980. https://doi.org/10.1016/j.biortech.2020.122980
- Kandler, O.: Carbohydrate metabolism in lactic acid bacteria. Antonie van Leeuwenhoek. 49, 209–224 (1983). https://doi.org/10.1007/BF00399499
- Kandylis, P., Bekatorou, A., Pissaridi, K., Lappa, K., Dima, A., Kanellaki, M., Koutinas, A.A.: Acidogenesis of cellulosic hydrolysates for new generation biofuels. Biomass Bioenerg. 91, 210–216 (2016). https://doi.org/10.1016/j.biombioe.2016.05.006
- German, D.. Jojoa-Unigarro, Simón González-Martínez: (2021) Kinetic analysis of the methanization of the byproducts from OFMSW fermentation. J. Chem. Technol. Biotechnol. Published on line for the first time, September 2021. https://doi.org/10.1002/jctb.6921. ISSN 0268–2575
- Figueroa-Escamilla, L., González-Martínez, S., Campuzano, R., Valdez-Vazquez, I.: Methane production and bromatological characteristics of the different fractions of organic municipal solid waste. Detritus. 15, 13–23 (2021). https://doi.org/10.31025/2611-4135/2021.15095
- Bouallagui, H., Torrijos, M., Godon, J.J., Moletta, R., Cheikh, R., Ben, Touhami, Y., Delgenes, J.P., Hamdi, M.: Two-phases anaerobic digestion of fruit and vegetable wastes: bioreactors performance. Biochem. Eng. J. 21, 193–197 (2004). https://doi.org/10.1016/j.bej.2004.05.001
- Rajeshwari, K.V., Lata, K., Pant, D.C., Kishore, V.V.N.: A novel process using enhanced acidification and a UASB reactor for biomethanation of vegetable market waste. Waste Manag. Res. 19, 292–300 (2001). https://doi.org/10.1177/0734242X0101900405
- Baldi, F., Pecorini, I., Iannelli, R.: Comparison of single-stage and two-stage anaerobic co-digestion of food waste and activated sludge for hydrogen and methane production. Renew. Energy. 143, 1755–1765 (2019). https://doi.org/10.1016/j.renene.2019.05.122
- Campuzano, R., González-Martínez, S.: Characteristics of the organic fraction of municipal solid waste and methane production: A review. Waste Manag. 54, 3–12 (2016). https://doi.org/10.1016/j.wasman.2016.05.016
- Ma, X., Yu, M., Song, N., Xu, B., Gao, M., Wu, C., Wang, Q.: Effect of ethanol pre-fermentation on organic load rate and stability of semi-continuous anaerobic digestion of food waste. Bioresour Technol. J. 299, 122587 (2020). https://doi.org/10.1016/j.biortech.2019.122587
- Valentino, F., Munarin, G., Biasiolo, M., Cavinato, C., Bolzonella, D., Pavan, P.: Enhancing volatile fatty acids (VFA) production from food waste in a two-phases pilot-scale anaerobic digestion process. J. Environ. Chem. Eng. 9, 106062 (2021). https://doi.org/10.1016/j.jece.2021.106062
- Schievano, A., Tenca, A., Lonati, S., Manzini, E., Adani, F.: Can two-stages instead of one-stage anaerobic digestion really increase energy recovery from biomass? Appl. Energy. 124, 335–342 (2014). https://doi.org/10.1016/j.apenergy.2014.03.024
- Yu, M., Gao, M., Wang, L., Ren, Y., Wu, C., Ma, H., Wang, Q.: Kinetic modelling and synergistic impact evaluation for the anaerobic co-digestion of distillers' grains and food waste by ethanol pre-fermentation. Environ. Sci. Pollut Res. 25, 30281–30291 (2018). https://doi.org/10.1007/s11356-018-3027-6
- Chu, D., Barnes, D.J.: 2016. The lag-phase during diauxic growth is a trade-off between fast adaptation and high growth rate. Scientific Reports 6, article number 25191, 1–15. https://doi.org/10.1038/srep25191
Table 1. Concentrations of metabolites at the end of the 6-day fermentation. All values are concentrations in mMol/L. NOM, naturally occurring microorganisms.
|
|
Lactic acid |
Ethanol |
Acetic acid |
|
Fermentation with NOM |
132±4 |
51±6 |
30±5 |
|
Fermentation with yeast + NOM |
98±15 |
108±10 |
32±4 |
|
Fermentation with UASB sludge +NOM |
178±11 |
22±3 |
28±2 |
Table 2. COD, solids, and hydrolysis rate at the beginning and end of the 6-days fermentation. NOM, naturally occurring microorganisms.
|
Inoculum |
COD (g/L) |
Solids (g/L) |
Hydrolysis rate |
|||||||
|
Initial |
Final |
Initial |
Final |
(gCODdiss/ kgTS·d) |
(gCODdiss/ kgCODt·d) |
|||||
|
Total |
Diss. |
Total |
Diss. |
TS |
VS |
TS |
VS |
|||
|
NOM |
75±10 |
23±3 |
64±6 |
25±2 |
52±4 |
40±4 |
41±5 |
30±2 |
35 |
24 |
|
Yeast+NOM |
82±6 |
25±1 |
74±7 |
29±2 |
54±5 |
42±4 |
52±4 |
34±3 |
66 |
44 |
|
UASB sludge+NOM |
74±3 |
22±1 |
68±6 |
23±1 |
54±6 |
42±4 |
52±2 |
38±1 |
12 |
9 |
Table 3. Specific methane production for the three cases plus unfermented OFMSW as reference. Included are also the correlation coefficients and the intercept values obtained from Figure 3.
|
|
Slope (Specific CH4 production, NmL/gVS) |
Intercept |
Corr. Coef. |
|
Unfermented OFMSW |
220 |
235 |
0.9639 |
|
Fermented with NOM |
322 |
150 |
0.972 |
|
Fermented with yeast+NOM |
420* |
NA |
NA |
|
Fermented with UASB sludge+NOM |
244 |
209 |
0.9771 |
*Obtained with two values. NA, not available
Table 4. Comparison of the specific methane production and conditions of fermented organic solid wastes.
|
Author |
Substrate |
Inoculum for fermentation |
Methanogenesis |
CH4 production (NL/kgVS) |
|
This research |
OFMSW with garden trimmings |
Yeast. Batch. 35°C. pH 4. SRT 24 h. |
Batch, 35˚C, reaction 30 d |
62 |
|
UASB sludge. Batch. 35°C. pH 4. SRT 24 h. |
Batch, 35˚C, reaction 30 d |
101 |
||
|
Native microorganisms. Batch. 35°C. pH 4. SRT 24 h. |
Batch, 35˚C, reaction 30 d |
196 |
||
|
Without fermentation |
Batch, 35˚C, reaction 30 d |
217 |
||
|
Ma et al., 2020 |
Food waste |
Yeast. Batch. SRT 24 h, 35˚C |
Semicontinuous, 37˚C, HRT 20 d, OLR 5.5 gVS/L·d |
203 |
|
Without fermentation |
Semicontinuous, 37˚C, HRT 20 d, OLR 5.5 gVS/L·d |
184 |
||
|
Yu et al., 2018 |
Food waste and distiller's grains |
Yeast. Batch. SRT 24 h. 35˚C |
Batch, 35˚C, Inoculum/substrate ratio 3 (VS), digestion time 38 days |
280 |
|
Ganesh et al., 2014 |
Fruit and vegetable waste |
Mixed cultures for VFA Batch. SRT 5 to 10 d. |
Methanization only of the liquid fraction. CSTR, HRT 30 d, 37˚C, OLR 7.0 gVS/L·d |
300 |
|
Bouallagui et al., 2004 |
Fruit and vegetable waste |
Mixed cultures for VFA Reaction 21 h. 35˚C |
Methanization only of the liquid fraction. Batch, HRT 21 h, 35˚C, OL 1.29 gDQO/L·d, SRT 10 d |
320 NL/kgCOD |
|
Rajeshwari et al., 2001 |
Vegetable market waste |
Mixed cultures for VFA Reaction 40 h. 35˚C |
Methanization only of the liquid fraction. UASB, HRT 16 h, 37˚C, OL 19.6 gDQO/L·d |
350 |
|
Shen et al., 2013 |
Fruit and vegetable waste |
Mixed cultures for VFA CSTR. HRT 10 d. 37˚C |
HRT 10 d, 37˚C, OLR 4.0 gVS/L·d |
351 |
|
Without fermentation |
HRT 30 d, 37˚C, OLR 4.0 gVS/L·d |
328 |
||
|
Baldi et al., 2019 |
Food waste |
Native microorganisms. CSTR. HRT 3 d, 35˚C. OLR 14.2 kgVS/m3·d |
CSTR, HRT 12.8 d, 35˚C, OLR 2.5 kgVS/m3·d |
705 |
|
Without fermentation |
CSTR, HRT 17 d, 35˚C, OLR 2.5 kgVS/m3·d |
694 |
Table 5. Specific methane production and values were obtained with the Gompertz model.
|
Inoculum for fermentation |
Substrate concentration (gVS/L) |
CH4 yield (N.L./kgVS) |
Values obtained with Gompertz |
|||
|
Pmax (NL/kgVS) |
Rmax (NL/kgVS·d) |
λ (d) |
R2 |
|||
|
NOM |
1.25 |
707 |
710,4 |
114,4 |
-4,7 |
0,9980 |
|
2.5 |
569 |
604,2 |
64,0 |
-6,5 |
0,9937 |
|
|
5 |
389 |
441,6 |
29,4 |
-10,5 |
0,9846 |
|
|
10 |
270 |
301,5 |
33,0 |
4,8 |
0,9802 |
|
|
Yeast + NOM |
1.25 |
664 |
667,0 |
110,1 |
-3,9 |
0,9980 |
|
2.5 |
547 |
567,5 |
64,1 |
-5,6 |
0,9939 |
|
|
5 |
195 |
223,2 |
16,1 |
-8,2 |
0,9951 |
|
|
10 |
157 |
NA |
NA |
NA |
NA |
|
|
UASB sludge + NOM |
1.25 |
677 |
696,7 |
92,5 |
-4,5 |
0,9982 |
|
2.5 |
486 |
503,2 |
64,1 |
-5,4 |
0,9964 |
|
|
5 |
344 |
471,1 |
26,3 |
-11,5 |
0,9890 |
|
|
10 |
180 |
268,2 |
20,7 |
3,2 |
0,9812 |
|
|
Raw OFMSW |
1.25 |
663 |
684,6 |
89,5 |
-3,5 |
0,9981 |
|
2.5 |
422 |
428,5 |
76,7 |
-3,4 |
0,9920 |
|
|
5 |
387 |
417,2 |
45,3 |
-6,1 |
0,9922 |
|
|
10 |
271 |
383,5 |
21,5 |
-8,1 |
0,9933 |
|
NOM: Naturally occurring microorganisms. NA, not available
{kind=link}