Chikungunya was first identified from southern Tanzania in 1952. It is characterized by abrupt fever mostly accompanied by joint pain. The symptoms may continue for a few days and occasionally stay for weeks, months, or even years. Additional manifestations comprise muscular pain, swelling at joints, nausea, headache, rash, and fatigue [1]. Chikungunya virus (CHIKV), the causal agent of chikungunya, is a member of the family Togaviridae and belongs to the genus Alphavirus. About 50 years of its initial isolation, CHIKV caused occasional outbreaks in Africa and Asia. Chikungunya re-emerged in the first decade of the present century in the Indian subcontinent, Caribbean islands, and French Guiana [2]. During 2004–2006, five hundred thousand people were reported infected from Kenya and surrounding locations. Subsequently, it spread to Indonesia, Maldives, Sri Lanka, Myanmar, and Thailand due to viremic travelers. In 2007, one hundred ninety-seven cases were reported from Italy in Europe. In 2014, Europe had the highest chikungunya burden, with approximately 1500 reported cases. In 2015 and 2016, 37480 and 146914 confirmed cases were reported from the United States. Pakistan had 8387 instances, whereas India had 62000. Chikungunya episodes have also been confirmed in Sudan, Yemen, Cambodia, and Chad more recently [3].
The vectors to carry the virus from viremic to native hosts were identified as mosquitoes Aedes aegypti and A. albopictus, recognized to be the most anthropophilic mosquitoes with a tight linkage to humans. After getting infected, they transmit the virus to humans through their bites, right into the bloodstream. This virus is an enveloped, icosahedral entity with a diameter of 65-70nm. Its genome is composed of positive-sense ssRNA of approximately 11.8 Kb. There are two open reading frames (ORFs), one of them coding for nonstructural proteins starting translation just after the entry into a host cell. This results in the synthesis of nonstructural polyprotein precursor nsP1-4. The ns polyprotein precursor was further processed by proteolytic cleavage into constituent nonstructural proteins nsP1, nsP2, nsP3, and nsP4 by virus-encoded nsP2 cysteine protease (nsP2pro). Replication of viral genome and subgenomic expression depend on these nsPs (nonstructural proteins). The nsP2pro is a multifunctional two-domain protein. The N-terminal domain is responsible for RNA helicase, RNA dependent 5'-triphosphatase, and nucleoside triphosphatase activities, whereas the C-terminal domain is liable for polyprotein cleavage into constituent nsPs [4, 5]. Cys478-His548 is the catalytic dyad of nsP2pro. Asn476, Cys478, Asn547, and Tyr544 are the main putative binding residues, while Met707, Asp711, and Leu670 as well play a significant role in substrate binding and identification. Trp549 is the most conserved residue in alphaviruses. Since the C-terminal domain of nsP2 is pivotal for the viral replication cycle, it is a tempting target for drugs and inhibitors [6]. In addition, nsP2 protease enters into the nucleus of the host cell and stops the expression of antiviral genes[7, 8]. To date, there is no vaccine or potent drug against CHIKV.
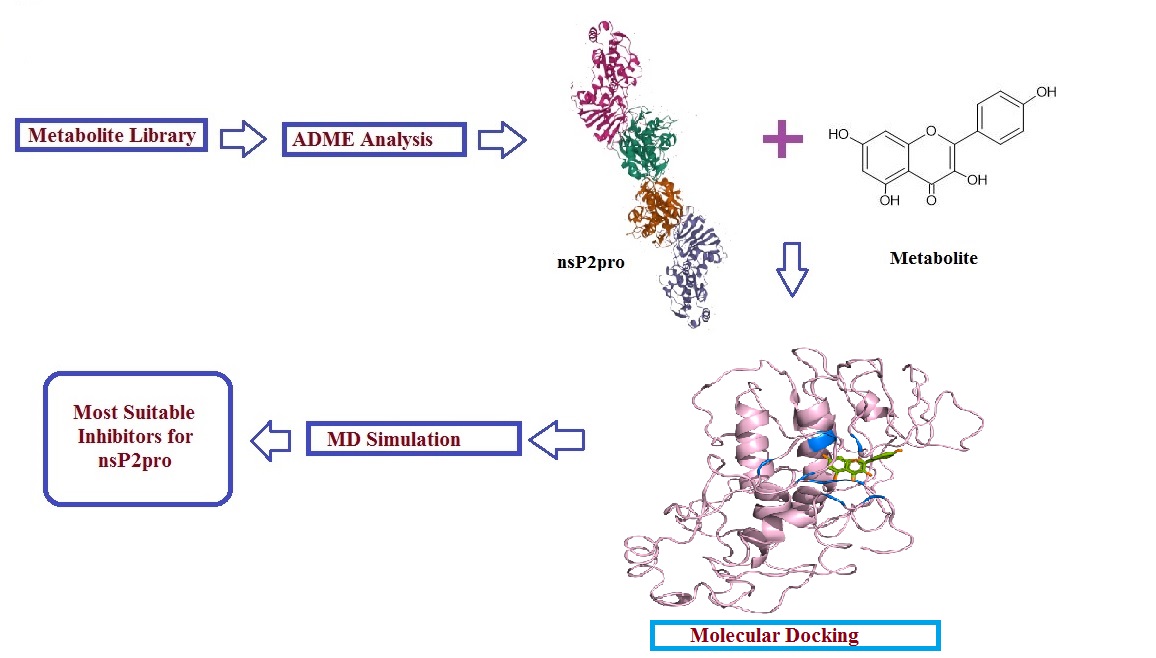
Several herbs have been identified to possess unique antiviral, antibacterial, antifungal and anti-cancer properties [9, 10]. Nigella sativa L. (black cumin or black seed), widely used as spice and medicine in various systems of medicines since ancient times [11], is also reported as “a cure for every disease except death” [12]. It is indigenous to North Africa, southern Europe, south and southwest Asia [9] with several bioactive metabolites. The oil extracted from black cumin seeds suppressed H9N2 (influenza virus) pathogenicity and improved immunity in poultry [13]. N. sativa seeds exhibited antiviral effects against Laryngotrachietis Virus (ILTV) in Chicken Embryo Rough Cells [14]. The diabetic patients infected with hepatitis C virus when treated with N. sativa were found with lower viral load and improved glycemic control [15]. When used against HIV findings confirmed the efficacy and potency of N. sativa seeds as potent anti-HIV [16] agents. This led to screening the bioactive metabolites against CHIKV using in silico approach. In the present study, we carried out molecular docking, atomistic explicit solvent MD simulations (molecular dynamic simulations), and MMPBSA (Molecular Mechanics Poisson Boltzmann Surface Area) based free energy calculations to investigate the potential inhibiting efficiency of selected bioactive metabolites from N. sativa to the nsP2pro, an important enzyme that participates in processing of nonstructural polyprotein complex. High binding scores, stability of the nsP2pro-chosen bioactive metabolite complexes, and high binding free energy values indicate that bioactive metabolites can be effective inhibitors for the nsP2pro.
{kind=link}