Taxonomy
The term Tetrahymenosis covers diseases affecting a number of fishes, crustaceans, mollus, beetle, dragonfly, salmon, slug, midge larvae and freshwater mussel species that caused by ciliates of the genus Tetrahymena which constitutes an abundant group inhabit various aquaculture and natural habitats (Table 1). Tetrahymena belongs to the phylum Ciliophora, Class Oligohymenophorea, order Tetrahymenida and family Tetrahymenidae [4]. Currently, the genus Tetrahymena Furgason, 1940 (Ciliophora, Hymenostomatida) is comprised of over 40 species [2–5, 13, 19]. Tetrahymena is generally pyriform-shaped, uniformly covered with meridional kineties, the buccal cavity is equipped with one curved undulating membrane with paired basal bodies organized in zigzag pattern (i.e. stichodyad), located on right edge of the buccal cavity, and three parallel membranelles. Within species, size variations depend on culture medium, physiological state and life stage (Corliss, 1973). The life cycle of Tetrahymena species is typified by the following developmental stages: theront, trophont, tomont and tomite.
Diversity of parasitic Tetrahymena
Tetrahymena species commonly exhibits a whole range of existence from the completely free-living state through stages of facultative parasitism to obligate endoparasitism [6]. Corliss (1971a) classified Tetrahymena parasitism into three types: facultatively parasitic forms (generally found free-living in nature but for which a parasitic existence is not uncommon), facultatively free-living forms (typically found in association with a host but capable in nature as well as experimentally) and parasitic forms (as endoparasites associated with specific hosts). To date, about ten Tetrahymena species (T. corlissi, T. pyriformis, T. stegomyiae, T. chironomi, T. vorax, T. rostrata, T. limacis, T. rotunda, T. glochidiophila and T. papula; Table 1) have been generally reported as pathogenicity to various invertebrates and vertebrates [16–18, 22].
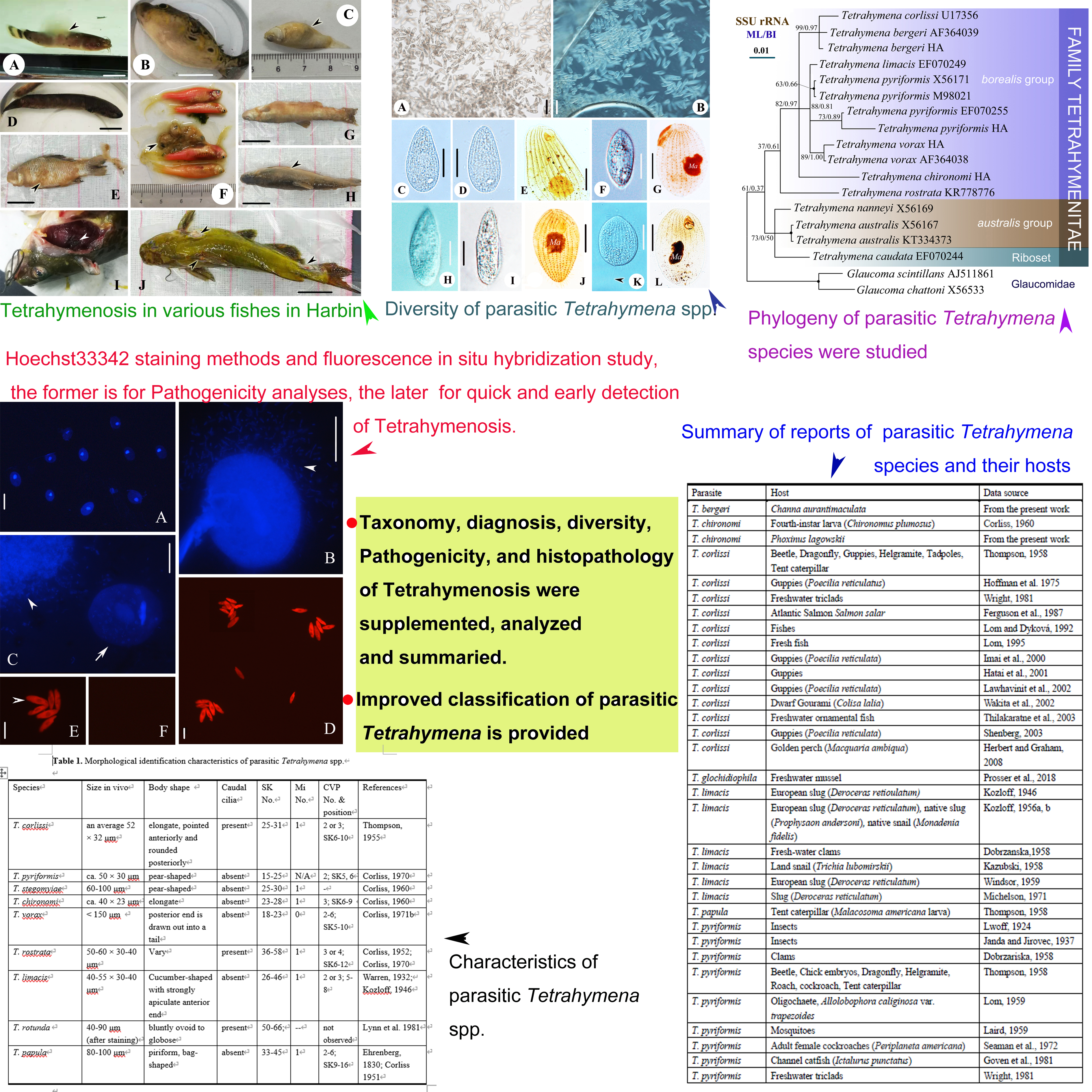
In the current work, T. pyriformis, T. vorax, T. chironomi and T. bergeri are the four species isolated from fish farms or markets in Harbin. The cells commonly showed a high densities and vitalities when just isolated from the various fishes (Figs 1, 2). The detailed morphology of the four species are reported before, and the Harbin populations of the four species are similar with those previous descriptions (Figure 2) [1, 3, 7, 42]. The topologies of BI trees are similar to that of ML trees, so only the ML trees are shown in Figure 4 with branch support values for both analyses. In the consensus topology, all the parasitic Tetrahymena spp. (Table 1) having sequence data are selected to constructed trees. As is shown, all the parasitic forms cluster in the ‘borealis’ group. The clustering pattern indicates that ‘borealis” Tetrahymena spp. might have a greater probability to become parasitism [13, 43]. In addition, Harbin population of T. pyriformis, T. vorax, and T. bergeri cluster with their previous populations, respectively, which also supports the morphological identifications.
Diagnostic features:
Pathogenicity/Histopathology.
As pathogenic parasites, histopathology of parasitic Tetrahymena spp. have been studied in detail and reported many times [16, 18, 22, 44–48]: possessing typical whitish lesions on the body surface, and affected organs include the skin, musculature, viscera, eye socket and spinal cord; masses of ciliates can be detected in copious amount of mucus and between spaces in the damaged tissues. In fishes, extensive studies reporting the organ-specific pathological changes when infected with Tetrahymenosis are available, and typical gross lesions of Tetrahymena infection are characterized by protrusion of scales, swelling, ulcerative wounds on the skin, blindness, and protrusion of eyes [47, 48, 22, 18]. In our recent study, four Tetrahymena species were isolated Malapterurus electricus, Tetraodon palembangensis, Poecilia latipinna, Channa aurantimaculata, Carassius auratus, zebrafish, Phoxinus lagowskii, Pelteobagrus fulvidrac from farms in Harbin. At arrival, fish did not have any apparent gross signs of Tetrahymena infection. Nonetheless, the parasite was observed in fresh-mount preparations on the skin, in the gills and in internal organs (Figure 2). Noticeable, many cells of Tetrahymena bergeri were observed in the skin mucus of Phoxinus lagowskii, individuals of P. lagowskii exhibited symptoms that no feeding, staying upside down, gasping attack edges and secreting a lot of mucus (Figure 3G, H, arrowhead). Tetrahymena pyriformis were observed from guts of diseased zebrafish (Figure 3F, arrowhead). Pelteobagrus fulvidraco had many large, round deep skin lesions all over the body with hyperaemic margins exposing the skeletal muscle, and scrapings revealed the presence of T. vorax (Figure 2G, arrowheads). Gills of all diseased fish were shriveling, dark and anaemic, gills of several fish had dark-red ulceration (Figure 2I; arrowhead). Some cells of T. vorax were also detected in gaps among the gill filaments.
Skin lesions or mucus of fishes were directly stained in vivo by Hoechst33342 staining method, and Figure 3A–C show the process of facultatively parasitic Tetrahymena sp. eating bacteria around the cell (Figure 3B), and Tetrahymena sp. eating tissues (skin or gill) as well as bacteria of host (Figure 3C). Cytoplasm of Tetrahymena sp. always contain many (approximately 2–5 μm in diameter) bacteria-or fish tissue- filled food vacuoles and variable-sized granules, distributed randomly (Figure 3C, arrowhead). The result indicates that facultatively parasitic Tetrahymena sp. can physically devour tissue, not only by producing enzyme [49].
Detection and identification using FISH
The probe Tetr2020 evaluated with the probe match tool of the ARB software package and the GenBank BLAST tool showed that they are specific to Tetrahymena spp. The probe Tetr2020 (5’- TGTAGTAGCCGTTTCTCAG-3’) had at least two mismatches to other closely related species like Uronema spp., Pseudocohnilembus spp. and Cyclidium spp. FISH with the probes Tetr2020 resulted in the presence of a red fluorescence signal each for T. pyriformis, T. vorax, T. chironomi and T. bergeri (Figure 3D, E), clearly distinguishable from the faint autofluorescence signal achieved with negative-control hybridizations using the Tetr2020 probe to hybridize the untargeted ciliates Pseudocohnilembus persalinus, Uronema marinum and isolated from the same hosts (Figure 3F). Thus, the probes Tetr2020 is also a general probe suitable for widely and rapidly detecting Tetrahymenosis.
{kind=link}