FPS detects the binding of molecules in real-time through changes in the dye’s local environment (Häußermann et al., 2019). FPS, based on SwitchSENSE technology, relies on a biochip with covalently attached single stranded anchor DNA for target protein immobilization at a distance of approximately 30 nm (Knezevic et al., 2012), thereby potentially precluding re-binding and avidity effects. The peptide (analyte) binding is reported by a fluorescent reporter close to the immobilized protein of interest (ligand) (Figure 1 A) and consequently independent of unspecific binding of the analyte to the chip surface. Importantly, FPS neither requires direct fluorescent labelling of the ligand nor the analytes, thereby avoiding disturbance of their functional integrity or other dye-mediated artefacts. In contrast to other recently reported kinetic methods (Stein et al., 2021), FPS allows for the analysis of slow (<10-4 s-1) off-rates and fast on-rates (>106 M-1s-1).
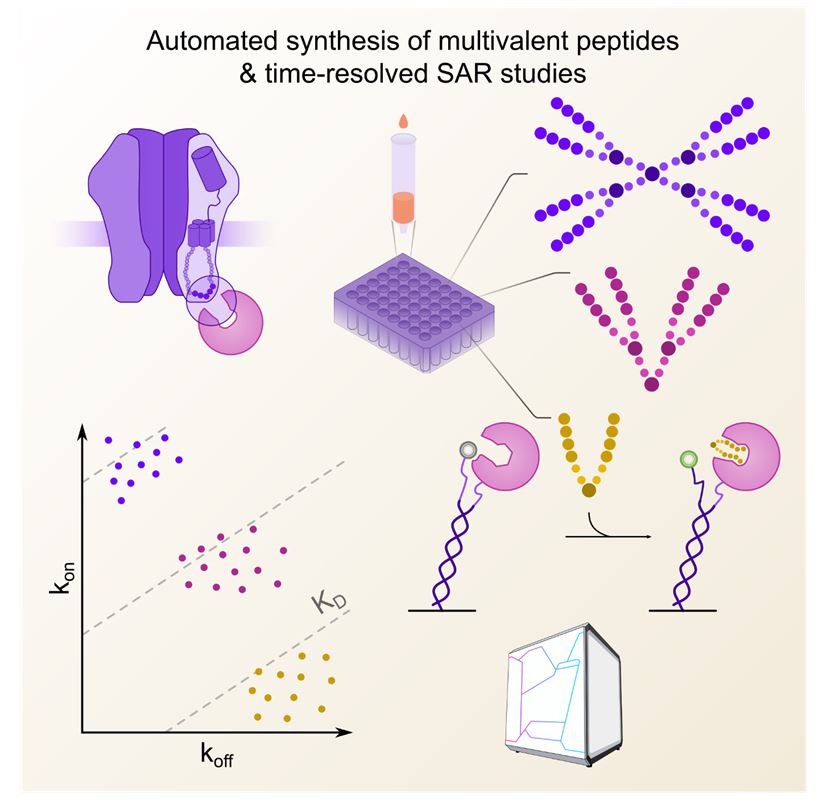
While the workflow is designed to be applicable to any multivalent system where combinatorial display is feasible, we here use the neuronal scaffolding protein gephyrin (Tyagarajan and Fritschy, 2014) (geph) and its structurally resolved (Maric et al., 2014) interactor, the glycine receptor (GlyR) β subunit. PPIs within receptor protein complexes (Rosenbaum et al., 2020) and specifically scaffolds of the neuronal synapses are explored with multivalent chemical probes (Maric et al., 2017, Sainlos et al., 2009, Bach et al., 2012) and studied in pharmacological context (ClinicalTrials.gov, NCT04689035) (Schulte and Maric, 2021). Dimeric, tetrameric and octameric binders were synthesized using an accessible and broadly applicable strategy by combining binding sequences with Polyethylene glycol (PEG) linkers and L-Lysine cores (Nomizu et al., 1993) as branching points (Figure 1 A). Using varied geph binding sequences, PEG linkers of variable length and up to three branching points, we synthesized a total of +100 unique multimeric compounds (Supplementary Table 1), differing over one magnitude in molecular weight.
For the FPS measurements, the otherwise unlabelled receptor binding geph E-domain (gephE) was coupled to the ligand strand while a fluorescent reporter was attached to the dye strand (Figure 1 B). Among six tested dyes, the fluorescence change was highest for the green dye B (Dynamic Biosensors GmbH, DE) (Supplementary Figure 1) which was therefore used in all subsequent FPS measurements. The functionality of this setup was demonstrated by recapitulating the structurally resolved geph-binding site of GlyRβ (398FSIVG402) (Maric et al., 2014) using a 1 µM library of unmodified, overlapping dimeric peptides with an offset of one amino acid (Figure 1 C).
Comparison of FPS with ITC, BLI and TRIC
Next, we assessed the reliability of apparent KD values determined in FPS by comparing this setup with commonly used immobilization- and in-solution-based PPI-quantification methods. Namely, real-time binding quantification using biolayer interferometry (BLI), HT temperature related intensity change (TRIC) quantification as well as precise calorimetric measurements (ITC) (Figure 2 A). Compared to ITC measurements, which can be considered the gold standard as they quantify directly and label-free in solution (Figure 2A), HT quasi label-free TRIC measurements (Figure 2 B) recapitulate the same trend. The only exception being compound e=8, o1=0, o2=4, which was outside of the dynamic range. The BLI measurements (Figure 2 C) necessitated loading densities and ligand concentrations that did for effective dissociation of tetramers (Supplementary Figure 2) and octamers (Supplementary Figure 3). Thus, affinities could not be derived from single curves but were instead assessed through steady-state BLI measurements using multiple peptide concentrations. The determined KD values only partly recapitulated the affinities determined in ITC, possibly due to avidity effects such as re-binding.
Along the same line, BLI overestimated the affinity of the tetramers and further enabled the measurement of e=5, o=2, a lower affinity dimer. Conversely, the on- and off-rates of dimeric peptides were resolvable in BLI (Supplementary Figure 4). However, a poor signal-to-noise ratio (SNR) was observed for small dimeric peptides (Supplementary Figure 4 C and D). In stark contrast, FPS enabled measurements of dimeric, and tetrameric compounds independent of compound size (Figure 2 D). The resolved binding hierarchy is in line with ITC and TRIC, similar so the apparent dynamic range.
Next, we compared the protein sample consumption of the four different biophysical PPI quantification methods (Figure 2 E). In terms of target protein consumption by weight, FPS performed second best among the methods employed, consuming 28.5-fold less protein than BLI measurements for sensor functionalization (0.64 µg for one FPS sensor chip versus 18.25 µg for 8 BLI biosensors), 285-fold less than ITC (182.4 µg for one run) and 2.2-fold more than TRIC (0.29 µg for a 16-point dose response) (Figure 2 B).
To facilitate the determination of kinetic binding parameters of hundreds of peptides with a short turnover, we explored the possibility to directly couple FPS to low µM scaled solid-phase peptide synthesis. Consequently, we determined the intra-synthesis reproducibility of real-time affinity measurements of multimeric, unmodified peptides in FPS. KD values and kinetic parameters could be determined with low deviation using independently synthesized dimers and tetramers (Supplementary Figure 5), indicating that the combined setup allows for reproducible and precise kinetic interactions studies.
HT determination of protein affinities and kinetics using FPS
Next, we used the established FPS setup to resolve the relationship between multimeric peptide architecture and binding kinetics. Specifically, an array of dimeric, tetrameric, and octameric compounds was subjected to FPS measurements at a fixed concentration of 1 µM to achieve sufficient signal amplitude for weaker binders (Figure 3). In addition to the on- and off-rates determined from functions fit to the obtained curves, association levels, at which the measured curves plateaued, were determined for each peptide. Overall, a prominent gain in affinity could be observed from dimers (Figure 3 A, low µM) to tetramers (Figure 3 B, high nM) and finally octamers (Figure 3 C, mid/low nM). Indeed, plotting of the obtained on-rates against the off-rates for each compound in a rate-map (Figure 3 D) reveals that multimer affinity primarily depends on the valency. This is in line with previous studies (Errington et al., 2019) which demonstrated that increased valency also increases the ability to create additional binding conformations within the configurational network. The second most important factor is the length of the epitope. This trend recapitulates the changes in binding strength that have been observed for the respective monovalent counterparts (Maric et al 2015). In the here studied multivalent system, the observed affinity gain is primarily driven by on-rate effects which vary over two magnitudes, while the off-rates vary only 5-fold across all tested species. Together, these data confirm the importance of the binding affinity of the single binding epitopes for higher valency systems, demonstrating the importance of on-rate effects.
FPS correlates multivalent topology and binding dynamics
To resolve how topological multimeric features determine on- and off-rates, our measurements included a series of compounds identical in epitope length and number but systematically varied scaffold arrangement. Plotting the obtained on-rates against the off-rates for each compound as a rate-map, together with color-coding of the topological adjustments visualizes a clear trend (Figure 3 E). The octamer with the lowest affinity is characterized by a multivalent architecture that enables flexible movement of the two sides of the multimer but sterically restricted movement of the epitopes themselves within the two tetramers. Vice versa, the multimeric architecture that enabled the greatest flexibility close to the epitopes while at the same time enforcing pre-orientation of the epitopes through sterical constrains in the centre displayed the highest affinity. The difference in affinity between both compounds is primarily driven by on-rate (4.5-fold) but also off-rate effects (1.4-fold).This dataset resolves the structure-activity relationship of multivalent geph-binders and provides a framework for the development high-valency, ultra-high affinity interactors in general.
Prediction of multivalent binding parameters
The 40 successfully measured compounds constitute only a small fraction of the theoretical possible combinations. To discern whether the obtained dataset allows to predict multimer properties, we used machine learning. Specifically, we applied the Random Forest Regressor using the encoded amino acids and analogous building blocks as training input. Here, the peptide sequences are represented through the amino acid composition (Spänig and Heider, 2019), which demonstrated overall good performance across multiple applications and provides easy interpretability (Spänig et al., 2021). First we explored whether the observed on- and off-rates and the resulting KD values can be reliably predicted. To this end, we applied a leave-one-out cross-validation and found a high correlation between predicted and observed KD values (Figure 4 A, Supplementary Table 2), off-rates (Figure 4 B) and on-rates (Figure 4 C) in case of the tetrameric and octameric group. In case of the dimeric peptides, a positive correlation was only found for the KD values. We additionally examined the correlation between observed association level and KD, on- and off-rate for each compound. Here, positive Pearson correlations were found in case of the dimeric group for KD (Figure 4 D) and especially off-rate (Figure 4 E) but not on-rate (Figure 4 F). In stark contrast, no or even negative correlations were found in the tetramer and octamer group when correlating the observed association level to the KD values (Figure 4 D), off- (Figure 4 E) and on-rates (Figure 4 F).
Taken together, these results indicate that for both lower avidity dimers and higher avidity tetramers and octamers, KD values can be reliably predicted across multivalent species using the outlined algorithm. In stark contrast, the association level may only be a representative metric for KD and off-rate for distinct topology classes.
Peptide binders with high avidity potently neutralize native gephyrin
Our FPS studies suggest that higher-order geph-binding multimers possess enhanced potency as inhibitors compared to their dimeric counterparts. Using a complementary peptide microarray-based approach (Schulte et al., 2021) with native geph from mouse brain lysates, we probed the geph neutralizing capacity of dimeric and tetrameric geph binders. Native geph was pre-incubated with dimeric and tetrameric binders (Figure 4 G) with varying architecture at increasing peptide competitor concentrations. Reduction in on-chip peptide binding by geph thus corresponds to neutralization of geph by competitor binding. Tetrameric binders exhibited up to two orders of magnitude more potent geph neutralization than the dimeric binders (Figure 4 H), thereby confirming the outcome of the FPS-based HT screen and further highlighting the value of the outlined approach for avidity-based binding optimization.
{kind=link}