SLAF based high-density mapping
A high-density and high-quality genetic map of goji berry assembled in the present study based on F1 population from 2 different species of Lycium through SLAF-seq strategy, demonstrated to be a highly cost-effective technique for the discovery of a large number of SNPs and wide-scale genotyping [34]. SLAF-seq has been utilized in many different crop species, even those without reference genome [40-35-24]. Gong et al. (2019) [24]; reported very first goji berry high-density genetic map based on intra-specific F1 population under ddRAD-seq and the current perpetual report on genetic mapping using SLAF-seq will further enhance the technology and improve understanding in molecular genetics of goji berry. Though, SLAF-seq approach has seldom been utilized in goji berry to construct a genetic map for QTL mapping. Comparatively, previous GBS based approaches such as RAD and ddRAD-seq have limitation due to technical and procedural complexity, cost unfeasibility and lack of pre-designed scheme and deep sequencing for ensuring accuracy and efficiency [40]. Instead, SLAF-seq, a high-throughput, cost-effective, deep sequencing technique based on double barcoding system accompanying a larger population and pre-designed strategy for large scale SNPs, InDel detection and genotyping [34].
Primarily, we projected a system on the basis of well-assembled reference genome sequence, due to the unavailability of goji genome, based on goji genetic distance and DNA content detail [37]; pepper was selected as reference genome to carry out the sequencing and ensure the uniformity, density and efficiency of markers production. Under the pilot experiment, RsaI and HinCII restriction enzymes were chosen and excised fragment (364-414bp) for sequencing. The SLAF library was evaluated by choosing the rice genome (Oryza sativa L. japonica) as a control for comparing with the reference genome to ensure reliability and validity. According to Sun et al. (2013) [34]; SLAF-seq defined pipeline, recommended quality scores were not less than Q30 and each individual minimal sequencing depth of 6-fold. Our dataset contained 3,021.32Mb paired-end reads and 95.04% reads were observed higher than or equal to Q30. The sequencing depth for parents was 66.43-fold and 15.23-fold for individuals, fulfilled the quantity and quality criteria of the markers, needed to conduct high-density genetic mapping. At last, we followed strict principles of filtering to get high-quality SLAFs (detail in methods). Totally, 214,961 polymorphic SLAFs were obtained with a polymorphism rate of 43.47%, higher than the previously identified [41, 42, 22]. In the current study, 8 segregation types were obtained with 24,329 maximum number of markers accounting for (aa×bb) homozygous group, which was greater than the previous reports [22,24]. Totally, 5,669 were obtained as high-quality markers following filtration process. All the demonstrated salient features represented markers accuracy, high-throughput, efficiency at a feasible cost, which further supports that this new SLAF-sequencing technique is highly recommended for utilization in goji berry.
High-density genetic map of goji berry
After establishing a successful SLAF library and the generation of high-quality markers. Only 3,495 SLAFs were integrated properly out of 5,669 on the genetic map along with 15,810 SNPs using High Map software, covering 1,649.03 cM on 12 linkage groups (LGs) of goji berry, with an average marker interval of 0.47 cM. The linkage map was sufficiently saturated with the smallest mean marker interval, as compared to the previous similar reports ranged from 0.48-0.95 cM [25-43-42]. A recent investigation by Zhao et al. (2019) [22]; on wolfberry genetic map based on 302 F1 individuals were constructed genetic map using 6,733 markers with a total genetic distance of 1702.45 cM and mean interval of 0.31 cM. The current linkage map contributed equal criteria to be highly dense with outstanding attributes than previously reported map [22-24]. Goji berry has gained popularity due to economic importance and its historical use as TCM [44]. However, molecular breeding and genetics aspects of goji berry still need to be unfolded. In the current study, 305 F1 individuals were selected for high-resolution strategy, whereas larger number of individuals could bring higher recombination events [45-22], and accelerate the accuracy of fine QTL positioning [46].
The reliability of the genetic map was determined by haplotype maps, heatmaps, maximum gaps, mean gaps<=5, integrity of the individuals on the map and segregation distortion (P < 0.001). The haplotype maps and heat maps generated for each linkage groups revealed SLAFs distributed in a regular manner on all 12 LGs and correctly ordered while the pair-wise recombination rates were found lower significantly among adjacent markers except LG10 & 6. Furthermore, the maximum gap and average gap<=5 was larger than the previous study [22]. LG1, LG4 and LG7 were found with varied marker density and maximum gaps greater than > 10 cM. Probably, the low marker density areas depict strongly homozygous regions of the goji genome, a similar trend observed by Ollitrault et al. (2012) [47]. Conversely, high-density marker areas depict centromeric location on the chromosomes contributing larger physical distance leading to lesser genetic distance. Correspondingly, high-density areas may interact with the genome at some portion due to interspecific heterozygosity as described by Lindner et al. (2000) [48], Ollitrault et al. (2012) [47]. The integrity of the individuals on the genetic map was 99.03%, which reflected true high-quality and consistency of the map. Meanwhile, only 18 (0.34%) segregation distortion (P < 0.001) markers were observed in LG11 & LG12, significantly lower than the previously reported [35-22]. The segregation distortion among markers result might be due to the distant relationship of the parents and gametic or zygotic selection [49].
Our high-quality and high-density genetic map were found accurately designed and highly saturated than the previous reports [33-22]. We claim that the linkage map is precise and exhibit high-throughput, which would provide insights on the molecular biology of economically important medicinal tree plant and subsequently strong grounds for QTL mapping analysis and candidate gene identification of particular fruit size-related traits in goji berry. Further, it will ensure to be an essential source for comparative genomic studies [59]; and aid in assembling the reference genome of goji berry.
QTL mapping analysis for fruit size-related traits
QTL mapping based on high-density genetic maps can provide useful estimation in specific quantitative traits and data mining [29]. For conducting QTL analysis, different software`s are available limited to the crossing models such as IciMapping [50], MapQTL [51], and so on. The current study is based on F1 interspecific population and cross-pollinated (CP) model, which is different than other population used for mappings such as RILs, back cross and double haploids [46]. Thus, MapQTL with composite Interval mapping (IM) is definitely a suitable software for conducting QTL analysis in CP model, besides it has been successfully utilized in different species, pepper [46], cucumber [43], and walnut [31]. Zhang et al. (2010) [52]; successfully plotted QTL mapping for 8 traits in sweet cherry. In a recent report by Gong et al. (2019) [24]; a total of 32 QTLs was found corresponding to six photosynthesis-related traits using MapQTL in L. barbarum L. Whereas, the interspecific high-density genetic map of goji berry has been explored for six agronomically important traits utilizing MapQTL [22].
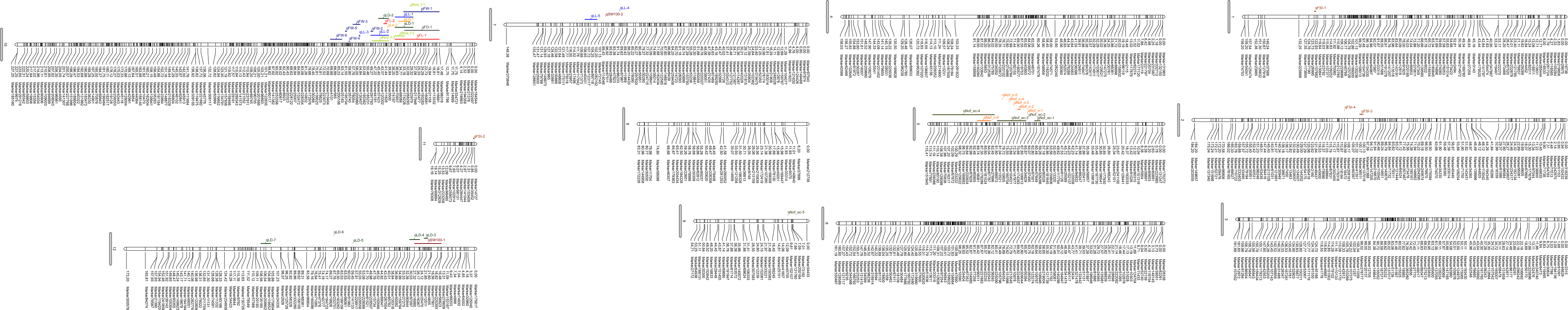
We employed MapQTL v. 6.0 and characterized a large set of QTLs controlling fruit size and quality-related traits. Particularly, 6 promising QTLs significantly observed on LG10 targeting FW, FL, LL, LD and LA, under 3 years consistently. While, 23 QTLs were detected in both years for FD, FSI, Nof/ec, Nof/n, Nos/f and 100SW (Fig. 3). Certainly, QTLs detected in 3 years dataset were considered stable. Fruit weight stable QTL qFW10-3.1 was detected on LG10 with LOD up to 8.17 and phenotypic variance 11.6%. Conversely, no fruit weight stable QTL was detected in previous wolfberry genetic map [22]. The fruit weight QTLs has been reported in different Solanaceae species, for example in tomato 28 FW QTLs were identified. Specifically, fw1.1, fw2.1, fw2.2, fw3.1, fw3.2 and fw11.3 major fruit weight QTLs mapped in almost four interspecific individual studies and accounted for fruit weight variation during tomato evolution [21]. Similarly, 5 fruit weight QTLs were detected in pepper, and fw3.2 was found with significant impact and larger phenotypic variance (R2=0.12-0.15)3. Pereira et al. (2018) [41]; investigated QTLs regarding fruit morphological traits in Cucumis melo L. and found FWQU5.1 was the major QTL on LG5 controlling fruit weight with maximum PVE of 28.3%. For fruit length QTL analysis, the current study detected the promising QTL, qFL10-2.1 on LG10 with a maximum of 8.88 LOD and 12.4% phenotypic variance. Similarly, Zhao et al. (2019) [22] detected 4 stable FL QTLs on LG11 harboring 16 markers with LOD and PVE up to 17.24 and 30.9%, respectively, in their interspecific genetic map of goji berry. Whereas several previous reports identified fruit length QTLs spanned on different chromosomes, and fruit diameter QTLs as well [53-25].
Among the leaf related traits, qLL10-2.1 stable QTL corresponding to leaf length was spanned on LG10, with peak LOD value and PVE up to 14.21 and 19.3%, respectively. While, qLD10-2.1 & qLD12-4.1, promising QTLs controlling leaf diameter located on LG10 and 12, with highest LOD and phenotypic variance of 7.17 & 8.1 and 10.3% & 11.5%, respectively. The stable QTL responsible for leaf area was qLA10-2.1 spanned on LG10, with a corresponding peak LOD of 7.74 and PVE 11%. Two stable QTLs reported regarding leaf index, qLI10-2 and qLI11-2 on LG10 and 11, harbored 4 and 20 markers, respectively. However, no stable QTLs controlling leaf length and leaf width were detected in previous goji berry genetic map [22]. The 2 leaf shape QTLs in pea were detected on LG7 with LOD values up to 2 [42]. In this study, the fruit length QTL, qFL10-2.1 was stable and colocalized with leaf related QTLs such as qLL10-2.1, qLD10-2.1, and qLA10-2.1 (Fig. S9a-o; Additional file 3). Meanwhile, the markers detected under qFL10-2.1 were also similar to leaves related stable QTLs. Remarkably, co-localization of traits linked QTLs frequently reflected a significant correlation level among different traits. Likely, fruit diameter and pericarp thickness analysis revealed 3 common positions on the genetic map of pepper (r=0.86, 0.87), reflected dual genomic regions with common interaction for both traits [3].
It is one assumption that QTLs detected on the same genomic region targeting various traits may occur due to pleiotropy or linkage phenomenon [3]. While considering promising QTLs, only 17 same markers were found associated with FL, LL, LD & LA. Another set of 5 markers is also worth mentioning, which were located in stable QTLs related to FL, LL & LA on LG10. Previously, in pepper, 3 markers were significantly related to several traits, for example, 1 marker (LG8) influenced six traits, another marker (LG2) associated with four traits and third marker (LG3) related with five different fruit development traits [3]. Therefore, we may speculate that those particular 5 genomic regions or QTLs may involve either a specific locus associated with pleiotropy, which might influence different aspects of goji fruit and plant architecture development or several linked loci indicates specific influence on plant developmental responses. Further studies will require to target these promising QTLs for determining the potential candidate genes influencing different fruit development traits in goji berry. The fruit sweetness (SSC) QTLs, qFS3-1 and qFS5-2 were detected on LG3 & 5 with relative LOD of 5.48 and 9.2% PVE and qFF10-1 responsible for fruit firmness were spanned on LG10 with peak LOD of 4.81 and PVE 8.1%. Ben Chaim et al. (2001) [3]; reported similar event by identifying 3 QTLs in each year but none as stable, corresponding to soluble solids concentration (SSC) in pepper with the largest phenotypic effect (R2=0.12). Whereas, 2 QTLs, fi9.1 & fi11.1 were detected controlling fruit firmness trait in pepper with 14% total variance [3]; which indicated consistency with our findings. Another study conducted a QTL mapping analysis in watermelon found QBRX2-1 and QBrix6 flesh sweetness QTLs [54]; harbored sugar transport gene ClTST2 [55].
However, transcriptome sequencing analysis is also essential to evaluate the expression pattern between different fruit and leaf developmental stages, which can exacerbate the mining process of candidate genes identification [43-22]. It has been revealed that promising orthologous QTLs or genes in similar crop species influence the same morphological or phenotypic impacts. In fact, some physiologically associated traits comparison has been done practically and identified similar genetic positions [56-3]. Before 90s, there was no QTL comparison has been made related to fruit-producing crops until the identification of fruit associated traits in tomato [4-21]. Ben Chaim et al. (2001) [3]; performed comparative QTL analysis between pepper and tomato and found quite interesting findings, regarding fruit shape and fruit weight traits. Exclusively, they observed orthologous QTLs in pepper and tomato targeting fruit weight on chromosomes 2 & 3 such as RFLP marker was found closely associated with fw2.1 and fw2.2 in pepper [3], and tomato [2], and another marker with fw3.2 and fw3.1 in pepper [3], and tomato [21], respectively. This perspective could be useful in the current study, like goji berry (Lycium spp.) relative to the same family. Thus, we might expect similar genetic identities or syntenic association as having been existed in tomato and pepper [3-57], tomato and eggplant [16], and tomato and potato [58]; which might be uncovered in later studies. In the current study, we have utilized common morphological traits related to the fruit and leaf development, which have a major contribution to the domestication of fruit crops [59]. To further evaluate the potential of goji population and explore additional QTLs, we already began developing recombinant inbred lines (RILs) by selecting full-sib mating with extremely distinguished offspring in 2019. RILs population have been successfully utilized in many crop species such as 146 RILs derived from intraspecific population employed for identification of first flower node QTLs [46], and similarly in peanut [39], melon fruit [41]. Additionally, enhanced QTL analysis could lead to exploring the genetic and molecular characterization of the agronomically important fruit traits in goji berry.

{kind=link}
{kind=link}
{kind=link}