Morphological traits variability analysis
A total of 11 morphological traits, including fruit weight (FW), fruit length (FL), fruit diameter (FD), fruit shape index (FSI), number of fruits per end cluster (Nof/ec), number of fruits per node (Nof/n), number of seeds per fruit (Nos/f), 100 seed weight (100SW), leaf length (LL), leaf diameter (LD) and leaf area (LA), were evaluated based on 2 individual years (2018-2019) and extra year (1819). The analysis of variance (ANOVA) of morphological traits of 2 individual years showed significant differences (P < 0.05 or P < 0.01), which indicated variation among 305 F1 progenies under different years (Table S1; Additional file 4). The descriptive statistical analysis showed coefficient of variation (CV%) ranged between 9 to 45 for FSI and Nof/ec in 2018, and 8 to 54 for FSI, FD and Nof/ec in 2019. However, 1819 data showed that CV% varied from 7 to 39 for FD and Nof/ec (Table S2; Additional file 5). The frequency distribution histogram and box chart among all morphological traits for individual years (2018-2019) and extra year (1819) showed normal distribution (Fig. S1a-z; Additional file 1). Normality test was performed using Kolmogorov-Smirnov (K-S) goodness of fit based on the absolute distance between cumulative distribution and values ranged from 0.02 to 0.11, suggested positive normal distribution of all traits evaluated. The correlation analysis (P < 0.05 or P < 0.01) revealed extreme significant positive association detected in a few comparisons among FL, FD, 100SW, Nos/f and FW; FD, FSI, 100SW, Nos/f, LL, LD, LA and FL; 100SW, Nos/f, LD, LA and FD; LL and FSI; LD, LA and 100SW; Nof/n, LL, LD, LA and Nos/f; Nof/ec, LL and Nof/n; LD, LA and LL; LA and LD or significant positive correlation between LD and FW; LL and FD; LD, LA and FSI; Nof/ec and Nos/f; LD and Nof/n. While highly significant negative correlation was detected between FSI and FD; Nof/n and 100SW; Nof/ec and 100SW (Table 1). The extremely positive correlated traits might indicate tight association among linked markers or even candidate genes due to pleiotropic effects, such as FW and FL, FD, Nos/f, and FL and FD, FSI, LL, LD, LA. This information could help determine candidate gene prediction.
Table 1 Combined correlation analysis among 11 agronomic attributes.
| |
FW
|
FL
|
FD
|
FSI
|
100SW
|
Nos/f
|
Nof/n
|
Nof/ec
|
LL
|
LD
|
LA
|
|
FW
|
1
|
|
|
|
|
|
|
|
|
|
|
|
FL
|
.657**
|
1
|
|
|
|
|
|
|
|
|
|
|
FD
|
.804**
|
.605**
|
1
|
|
|
|
|
|
|
|
|
|
FSI
|
0.051
|
.650**
|
-.204**
|
1
|
|
|
|
|
|
|
|
|
100SW
|
.342**
|
.320**
|
.312**
|
0.079
|
1
|
|
|
|
|
|
|
|
Nos/f
|
.317**
|
.238**
|
.263**
|
0.045
|
0.051
|
1
|
|
|
|
|
|
|
Nof/n
|
-0.055
|
-0.095
|
-0.043
|
-0.069
|
-.237**
|
.163**
|
1
|
|
|
|
|
|
Nof/ec
|
0.109
|
0.045
|
0.074
|
-0.003
|
-.152**
|
.125*
|
.520**
|
1
|
|
|
|
|
LL
|
0.099
|
.336**
|
.145*
|
.256**
|
0.047
|
.209**
|
.150**
|
0.112
|
1
|
|
|
|
LD
|
.144*
|
.297**
|
.228**
|
.134*
|
.172**
|
.276**
|
.122*
|
-0.006
|
.825**
|
1
|
|
|
LA
|
.156**
|
.261**
|
.174**
|
.133*
|
.200**
|
.262**
|
0.077
|
0.004
|
.821**
|
.887**
|
1
|
|
** Correlation is significant at the 0.01 level (2-tailed).
|
|
|
|
|
|
|
|
* Correlation is significant at the 0.05 level (2-tailed).
|
|
|
|
|
|
|
*FW, fruit weight; FL, fruit length; FD, fruit diameter; FSI, fruit shape index; Nof/ec, number of fruits/end cluster; Nof/n, number of fruits/node; Nos/f, number of seeds/fruit; 100SW, 100 seed weight; LL, leaf length; LD, leaf diameter; LA, leaf area.
Analysis of SLAF-seq data generated from F1 population
The sequencing data were examined to ensure the validity of the SLAF library construction. According to the goji genome size and GC content information [37], the pepper genome was selected as the reference genome utilizing self-developed predicted software and established protocol [6]. For optimal enzyme digestion the following guidelines were practiced such as; 1) lowest presence of selected digested fragments in the repetitive sequences, 2) regular distribution of digested fragments in the reference genome, 3) length and specificity of the digested fragment and specific pilot experiment system must comply uniformity, 4) the final number of digested fragments or SLAF tags must meet the expected number of tags [6, 70]. RsaI and HinCII were used for the SLAF library construction and 221,608 SLAF tags were obtained based on the pre-SLAF experiment.
The cross-check revealed normality of the SLAF library with paired-end mapping reads of 92.16% and enzymatic digestion efficiency of 93.11%. A total of 3,021.32M clean reads of data with a length of 200 bp were generated for goji berry, comprising 37M, 30M and 9M total paired-end reads in the female, male parents and offspring, respectively. The source data quality was ensured by average Q30 ratio of 95.04% and 39.62% GC. Correspondingly, in the female and male parent total number of SLAFs developed were 155,002 and 217,485 with an average depth of 72.43-fold and 60.43-fold of each SLAF marker, while for 305 F1 individuals 199,560 SLAFs were generated, with an average depth of 15.23-fold (Table 2). Of 494,472 high-quality SLAFs, 214,961 were polymorphic with 43.47% of polymorphism rate; furthermore, 279,356 non-polymorphic and 155 repetitive SLAFs were identified. After filtering out polymorphic SLAFs, a total of 40,616 SLAFs was classified into eight segregation patterns (Fig. 1). For the F1 cross-pollinated population, only 7 segregation patterns (ab×cd, ef×eg, hk×hk, lm×ll, nn×np, ab×cc, cc×ab) were retained for genetic mapping. We followed the genotyping quality criteria to filter out low quality and redundant markers using salient steps of filtering process (See “Methods” section). Collectively, we obtained 5,669 total SLAFs out of which only 3,495 high-quality SLAF markers were eventually utilized to construct an integrated (combined female and male parent map) high-density genetic map of goji berry with an average parental sequencing depth of more than 100-fold and offspring >10-fold (Table 3). Among 3,495 SLAF markers, ‘SNP_only’ were pre-dominant with 61.08% on the integrated map followed by ‘InDel_only’ (0.59%) and SNP & InDel (38.55%) (Fig. 2).
Table 2 Data description of SLAF-library Sequencing.
|
Samples
|
Total Reads
|
Q30 (%)
|
GC (%)
|
SLAFs
|
Total Depth
|
Average Depth
|
|
Female parent
|
37,527,272
|
94.86
|
40.69
|
155,002
|
11,227,106
|
72.43
|
|
Male parent
|
30,768,211
|
94.86
|
39.49
|
217,485
|
13,143,686
|
60.43
|
|
Offspring
|
9,682,038
|
95.04
|
39.62
|
199,560
|
3,038,879
|
15.23
|
|
Control
|
2,233,676
|
95.16
|
40.51
|
---
|
---
|
---
|
|
Total
|
3,021,316,939
|
95.04
|
39.62
|
---
|
---
|
---
|
Table 3 Detail Summary of high-quality SLAFs and SLAF markers
|
High-Quality SLAFs
|
|
|
Number of SLAFs
|
494,472
|
|
Average depth in female parent
|
72.43
|
|
Average depth in male parent
|
60.43
|
|
Average depth in individuals
|
15.23
|
|
Polymorphic SLAFs
|
|
|
Number of polymorphic SLAFs
|
214,961 (43.47%)
|
|
Number of non-polymorphic SLAFs
|
279,356 (56.50%)
|
|
Number of repetitive SLAFs
|
155 (0.03%)
|
|
High-Quality SLAF markers
|
|
|
Number of SLAF markers
|
3,495
|
|
Average depth in Female parent
|
111.85
|
|
Average depth in male parent
|
168.72
|
|
Average depth in individuals
|
35.80
|
*SLAF- Specific locus amplified fragment
Basic characteristics of the genetic map
After performing fine filtering process out of 5,669 SLAFs only 3,495 high-quality SLAF markers that met the quality standard were mapped onto the integrated genetic map comprised of 522 L. barbarum (male) and 3,143 L. chinense (female) markers with a ratio of 61.65%. (Fig. 3, Table 4). The average coverage of SLAF markers on the integrated genetic map was 111.82 cM for female and 168.72 cM male parent and 35.80 cM each F1 individual, respectively. The final map was 1,649.03 cM in length, confined to 12 LGs equal to the gametic chromosome number of L. barbarum with an average interval of 0.47 cM.
The largest and the most saturated linkage group was LG10, harbored 903 SLAF markers covering a length of 227.25 cM with the least average interval of 0.25 cM, while the smallest LG11 contained 36 markers with 19.16 cM genetic distance and 0.55 cM average interval. Surprisingly, the parental map largest linkage group was the same as that of the integrated map (LG10), harbored 142 markers spanning 147.16 cM for the male and 811 markers covering 292.32 cM for the female maps. However, LG8 and LG11 were short with an equal number of markers covering 20.47 cM and 2.15 cM genetic distances for the male map, respectively. In the same manner, LG9 belong to the female map was the smallest with 61.30 cM genetic distance (Table 4). The map uniformity revealed based on maximum gap and average gap <=5, which was experienced in LG1 with 16.99 cM and 99.10%, respectively. Moreover, we detected 15,810 SNPs ranges between 163 (LG11) to 4,013 SNPs (LG10) on the integrated map along with 6,135 transversions and 9,675 transitions (Table 4). Gong et al. (2019) [22], constructed the integrated genetic map based on intraspecific population with 23,967 SNP markers, spanned 964.03 cM of final genetic distance and 0.040 cM average marker interval, which reflected highly saturated map [24]. Similarly, another study reported wolfberry high-density SNP based genetic map with a final distance of 1702.45 cM and 0.31 cM average interval [22]. In comparison with previous reports, the current study revealed interspecific genetic map of goji berry with final genetic coverage of 1,649.03 cM and 0.47 cM mean marker interval. In particular, LG6 & LG10 was observed with least inter marker distances indicated maximum saturation and might be considered recombination hotspots in this population.
|
Table 4 Basic Characteristics of High-density genetic map of goji berry
|
|
|
|
|
|
|
| |
Number of markers
|
Distance (cM)
|
Average distance (cM)
|
Integrated map
|
SNPs
|
|
LGsID
|
Fm
|
Mm
|
Im
|
Fm
|
Mm
|
Im
|
Fm
|
Mm
|
Im
|
Gaps<=5%
|
Max. Gap
|
SNPs
|
Trv
|
Tri
|
|
LG1
|
269
|
79
|
335
|
170.32
|
127.48
|
159.2
|
0.64
|
1.63
|
0.48
|
99.1
|
16.99
|
1541
|
575
|
966
|
|
LG2
|
220
|
29
|
234
|
210.97
|
144.49
|
184.2
|
0.96
|
5.16
|
0.79
|
99.14
|
8.85
|
1044
|
411
|
633
|
|
LG3
|
335
|
49
|
364
|
181.81
|
139.5
|
162.4
|
0.54
|
2.91
|
0.45
|
100
|
4.64
|
1664
|
640
|
1024
|
|
LG4
|
373
|
30
|
391
|
170.51
|
111.96
|
158.91
|
0.46
|
3.86
|
0.41
|
99.23
|
11.2
|
1874
|
748
|
1126
|
|
LG5
|
228
|
45
|
259
|
125.24
|
103.94
|
115.74
|
0.55
|
2.36
|
0.45
|
100
|
4.94
|
1190
|
442
|
748
|
|
LG6
|
411
|
43
|
437
|
204
|
111.81
|
161.19
|
0.5
|
2.66
|
0.37
|
100
|
3.8
|
2028
|
773
|
1255
|
|
LG7
|
237
|
42
|
269
|
144.18
|
134.76
|
149.38
|
0.61
|
3.29
|
0.56
|
99.63
|
15.07
|
1195
|
476
|
719
|
|
LG8
|
46
|
6
|
50
|
87.06
|
20.47
|
83.31
|
1.93
|
4.09
|
1.7
|
91.84
|
6.2
|
219
|
89
|
130
|
|
LG9
|
31
|
11
|
39
|
61.3
|
37.23
|
55.09
|
2.04
|
3.72
|
1.45
|
94.74
|
7.01
|
170
|
64
|
106
|
|
LG10
|
811
|
142
|
903
|
292.32
|
147.16
|
227.25
|
0.36
|
1.04
|
0.25
|
99.89
|
7.62
|
4013
|
1581
|
2432
|
|
LG11
|
36
|
6
|
36
|
34.65
|
2.15
|
19.16
|
0.99
|
0.43
|
0.55
|
100
|
2.55
|
163
|
56
|
107
|
|
LG12
|
146
|
40
|
178
|
226.59
|
97.82
|
173.2
|
1.56
|
2.51
|
0.98
|
98.31
|
9.39
|
709
|
280
|
429
|
|
Total
|
3143
|
522
|
3495
|
1908.95
|
1178.77
|
1649.03
|
0.61
|
2.31
|
0.47
|
98.49
|
16.99
|
15810
|
6135
|
9675
|
|
*Fm= Female map, Mm= Male map, Im= Integrated map, Max.= maximum, SNPs= Single nucleotide polymorphism, Trv= Transversions, Tri= Transitions, LG=linkage group
|
Assessment of high-density genetic map of goji berry
To evaluate the quality of the genetic map, we performed several mapping approaches; Firstly, the integrity of all mapped markers was carried out between 305 F1 individuals to ensure the quality of the genotyping, accounting for 99.03% on average (Fig. S2; Additional file 1). Secondly, haplotype mapping was performed among each individual and both parents using 3,495 SLAFs to locate genotyping errors (Fig. S3a-l; Additional file 2). In the current study, the average percentage of the missing fragments were 0.0125%, represent the true quality of the map. It was verified that the interspecific F1 population was highly purified and suitable for high-density genetic map construction. DNA markers located on each linkage group were dispensed regularly with an average interval of 0.47 cM between adjacent markers despite high-rate of recombination events in F1 individuals. Thirdly, we displayed heatmaps to show the recombination frequencies among markers located on each linkage group to further evaluate the quality of the genetic map using pairwise recombination rate. The heatmaps depicted markers on the map were ordered accurately and pair-wise recombination rates were considerably low among adjacent markers and represented with yellow color diagonally. The purple color, particularly more visible in LG6 and LG10, reflected the highest recombination rates among adjacent markers (Fig. S4a-l; Additional file 2). Finally, 18 markers with segregation distortion (P < 0.001) were integrated in the construction of the genetic map, accounting for 0.34% of all mapped markers.
QTL mapping analysis
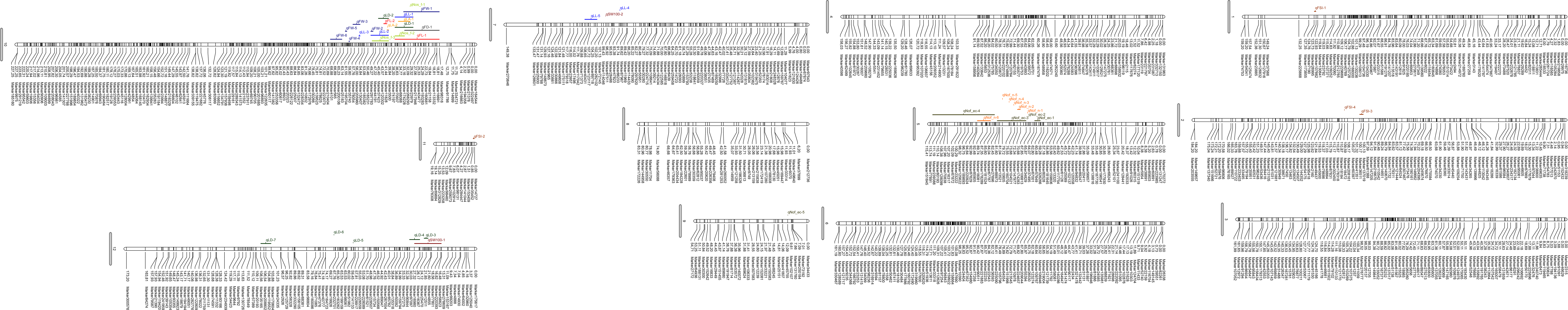
By using the high-density genetic map of goji berry, we plotted a large number of QTLs covering 13 traits. QTL analysis was performed using the composite interval mapping model with MapQTL v. 6.0 estimating 11 agronomical important traits for 2 individual years (2018-2019) and extra year (1819). Moreover, fruit quality traits; fruit sweetness (soluble solid content obrix) FS (SSC) and fruit firmness (FF) traits were examined during summer season, 2019. In order to set the LOD threshold, we performed 1000 permutation test and the threshold value was used to evaluate the statistical significance of each QTL. In this way, significant QTLs corresponding to particular traits was identified in 2 individual years and extra year consistently. As a result, most of the QTLs including promising QTLs relating to Nos/f, FD, FW, FL and leaf related traits were positioned in linkage group 10 (Fig. 4, Table 5). Other traits QTLs such as FSI, Nof/ec, Nof/n, 100SW, FS (SSC) & FF were located in different linkage groups (Fig. S5, S6, S7). In addition, qFS-1, qFS-2 corresponding to fruit sweetness with maximum phenotypic variance of 9.2% and qFF-1 to fruit firmness with 8.1% of PVE were mapped on LG3, 5 and 10, respectively (Fig. 4 and S6, Table 5). Collectively, 117 QTLs targeting 13 important traits were found (Table S3; Additional file 6). Of which 78 QTLs in 2 individual years with 6 stable QTLs (Table 5 and S3; Additional file 6) and 36 QTLs in extra year (1819) and only 3 QTLs in 2019 was detected. The number of markers located in corresponding QTLs ranged between 1 to 105, explaining highest PVE up to 19.3% with 14.21 Logarithm of Odds (LOD) covering all QTLs (Table 5 and S3; Additional file 6). QTLs detected corresponding to each trait in 2 individual years were described as follows:
Fruit weight (fw): Total 9 fruit weight QTLs were detected on LG10 and 6, of which major QTL was qFW10-1 with explained phenotypic variance of 11.1%. One stable QTL (qFW10-6.1) was located on LG10 with genetic distance from 65.89 to 71.86 cM and peak LOD value up to 6.01 along with corresponding PVE of 8.7% (Fig. 4). Moreover, 4 QTLs controlling FW was mapped on LG10 under extra year (1819) and qFW10-2 was confirmed as effective loci with maximum of 11.6% PVE (Table 5 and S3, additional file 6).
Fruit length (fl): 5 QTLs responsible for fruit length were found on LG10. The maximum phenotypic variance of 11.5%, corresponding to qFL10-1 considered as significant QTL. qFL10-2.1 targeting fruit length was observed as stable QTL on LG10, spanned an interval ranged from 44.64-56.88 cM with corresponding highest LOD of 6.54 and 9.5% phenotypic variance (Fig. 4). Another 4 FL QTLs were detected on LG10 in 1819 with highest PVE of 12.4% corresponding to qFL10-1 (Table 5 and S3, additional file 6).
Fruit diameter (fd): Only 5 QTLs corresponding to fruit diameter were located on LG10, spanned an interval ranged from 4.14-71.36 cM with a LOD value up to 7.8 and 11.1% PVE. qFD10-1 was detected as a major QTL for controlling fruit diameter with maximum phenotypic variance of 11.1% (Table S3, additional file 6). While 3 QTLs targeting FD were also found on LG10 with highest 12.8% PVE for qFD10-1 in 1819 (Table 5).
Fruit shape index (fsi): 8 QTLs controlling fruit shape index was observed on LG1, 2, 3, 7, 8 and 11, covered genetic interval ranged from 0.85-124.76 cM with LOD between 4.55 to 7.08 and explained variance of 6.6-10.2%. The significant QTL controlling fruit shape index was qFSI8-4 with maximum PVE of 10.2% (Table S3, additional file 6). In addition, 3 QTLs corresponding to FSI were detected on LG7 and 8 in extra year. The major QTL was qFSI8-3 with highest phenotypic variance of 8.5% (Table 5).
Number of fruits per end cluster (nof/ec): QTL analysis identified 8 QTLs targeting number of fruits per end cluster were positioned on LG5, 6, 8 and 9, spanned an interval ranged from 9.61-144.52 cM with LOD between 3 to 4.91 and 4.4 to 7.2% PVE. qNof/ec8-3 was detected as significant QTL with larger PVE of 7.2% (Table S3, additional file 6). In 1819 only 1 QTL was detected on LG5 with 8.1% PVE for Nof/ec (Table 5).
Number of fruits per node (nof/n): Total 7 QTLs corresponding to number of fruits per node were located on LG5 and 9, spanned an interval ranged from 14.05-92.48 with LOD value between 6.48 to 7.11 and 9.3 to 10.2% PVE. We detected qNof/n5-6 as major QTL with maximum 10.2% variance (Table S3, additional file 6). Only 2 QTLs targeting Nof/n were mapped on LG5 and 9 with extreme variation of 12.5% for qNof/n5-1 in extra year (1819) (Table 5).
Number of seeds per fruit (nos/f): We detected 4 QTLs on LG10, covered an interval ranged from 32.09-51.33 cM with LOD between 4.45 to 7.72 and PVE of 6.5 to 11%. Two major QTLs (qNos/f10-2, qNos/f10-3) were observed with the highest variance of 11% (Table S3, additional file 6). Another 3 Nos/f QTLs were identified on LG10 with larger PVE of 11.6% for qNos/f10-1 in 1819 (Table 5).
100 seed weight (100sw): 3 QTLs targeting 100 seed weight were detected on LG7, 9 and 12. These QTLs spanned an interval ranged from 14.05-100.07 with LOD value between 3.85 to 4.5 and 5.6 to 6.6% PVE. Q100SW7-2 was observed as a major QTL with maximum variance of 6.6% (Table S3, additional file 6). 7 QTLs targeting 100SW were also found on LG4, 10 and 12 with maximum PVE of 6.1% for q100SW10-1 in extra year (Table 5).
Leaf length (ll): 6 QTLs related to leaf length were mapped on LG7, LG10, while promising QTL (qLL10-2.1) positioned on LG10 starting from 42.77 to 51.91 cM interval bearing highest LOD of 10.48 with explained variance of 14.6% (Fig. 4). The major QTL (qLL10-2) was detected with maximum 14.6% PVE. Only 1 QTL (qLL10-1) was detected on LG10 with larger PVE of 19.3% in 1819 (Table 5 and S3, additional file 6).
Leaf diameter (ld): We observed 14 fruit diameter controlling QTLs were spanned on LG1, 10, 12, among them qLD10-2.1 and qLD12-4.1 represented stability throughout 2 individual years and positioned on LG10 and LG12 with covered distance ranged from 42.77-51.91 cM and 27.32-32.65 cM and corresponding highest LOD of 6.64 and 5.91 and explaining PVE% of 9.5 and 8.5, respectively ((Fig. 4). A qLD10-1 having maximum variance of 11%, considered as major QTL. Another 6 LD QTLs were located on LG1, 10 and 12 with highest PVE of 11.5% for qLD10-3 in extra year (Table 5 and S3, additional file 6).
Leaf area (la): 9 QTLs were found related to leaf area on LG1, 9, 10 and 12. Among them, qLA10-2.1 stable QTL targeting leaf area was detected on LG10 and covered genetic interval ranged from 39.84-58.12 cM with peak LOD of 7.74 and 11% explained variance (Fig. 4). Moreover, qLA10-1 was regarded as a major QTL based on maximum phenotypic variance of 12.5%. Only 2 QTLs corresponding to LA were found on LG10 with highest PVE of 5.4% for qLA10-2 in extra year (1819) (Table 5 and S3, additional file 6).
In our results, qLL10-1 was regarded as a major QTL with the largest explained 19.3% of variance and significant LOD of 14.21 (Table 5). Intriguingly, we encountered an interesting phenomenon of QTL co-localization among fruit and leaf related traits. Specifically, qFL10-2.1 was overlapped with tightly linked markers corresponding to LL, LD & LA among stable QTLs and represented a highly significant positive correlation (Table 1 and 5). The proportion of variance (PVE%) between promising QTLs was > 10%, which indicated major contribution of stable QTLs controlling corresponding important traits. Moreover, we had noticed a number of co-located QTLs such as fruit weight QTLs shared closely linked map positions with FL, FD, Nos/f, 100SW, LL, LD, LA and FF QTLs on LG10 consistently in 2 individuals year (2018-2019) and extra year (1819), which also indicated highly positive significant correlation as well (Fig. 4, Table 1 and 5).
Table 5 Promising QTLs detail information detected under 2 individual years and extra year (1819) data.
|
Traits
|
Year
|
QTLs
|
LGs
|
Interval (cM)
|
LOD
|
PVE (%)
|
| |
|
|
|
Start (cM)
|
End (cM)
|
distance (cM)
|
|
|
|
FW
|
2018/2019
|
qFW10-6.1
|
10
|
65.892
|
71.858
|
5.97
|
4.72-6.01
|
6.9-8.7
|
| |
1819
|
qFW10-1
|
10
|
17.483
|
34.771
|
17.29
|
6.51
|
9.4
|
| |
1819
|
qFW10-2
|
10
|
44.095
|
76.613
|
32.52
|
8.17
|
11.6
|
|
|
1819
|
qFW6-3
|
6
|
73.682
|
74.556
|
0.87
|
5.4
|
7.8
|
|
|
1819
|
qFW6-4
|
6
|
87.691
|
88.482
|
0.79
|
5.59
|
8.1
|
|
FL
|
2018/2019
|
qFL10-2.1
|
10
|
44.638
|
56.88
|
12.24
|
6.21-6.54
|
9-9.5
|
| |
1819
|
qFL10-1
|
10
|
13.739
|
51.909
|
38.17
|
8.88
|
12.4
|
| |
1819
|
qFL10-2
|
10
|
57.755
|
58.118
|
0.36
|
5.63
|
8.5
|
| |
1819
|
qFL10-3
|
10
|
59.799
|
59.799
|
0
|
5.87
|
8.5
|
|
|
1819
|
qFL10-4
|
10
|
62.134
|
62.134
|
0
|
5.86
|
8.5
|
|
FD
|
1819
|
qFD10-1
|
10
|
4.137
|
39.835
|
35.7
|
9.07
|
12.8
|
| |
1819
|
qFD10-2
|
10
|
65.892
|
73.183
|
7.29
|
7.43
|
10.6
|
| |
1819
|
qFD10-3
|
10
|
49.704
|
63.503
|
13.8
|
7.02
|
10.1
|
|
FSI
|
1819
|
qFSI7-1
|
7
|
104.247
|
108.987
|
4.74
|
5.51
|
8
|
| |
1819
|
qFSI7-2
|
7
|
109.885
|
111.761
|
1.88
|
5.45
|
7.9
|
| |
1819
|
qFSI8-3
|
8
|
60.56
|
65.763
|
5.2
|
5.89
|
8.5
|
|
Nof/ec
|
1819
|
qNof/ec5-1
|
5
|
67.85
|
69.165
|
1.32
|
5.56
|
8.1
|
|
Nof/n
|
1819
|
qNof/n5-1
|
5
|
61.076
|
92.992
|
31.92
|
8.86
|
12.5
|
| |
1819
|
qNof/n9-2
|
9
|
14.055
|
16.446
|
2.390
|
6.71
|
9.6
|
|
Nos/f
|
1819
|
qNos/f10-1
|
10
|
26.713
|
46.338
|
19.62
|
8.15
|
11.6
|
| |
1819
|
qNos/f10-2
|
10
|
48.095
|
48.095
|
0
|
6.22
|
9
|
| |
1819
|
qNos/f10-3
|
10
|
50.464
|
50.744
|
0.28
|
5.71
|
8.3
|
|
100SW
|
1819
|
q100SW10-1
|
10
|
69.787
|
72.522
|
2.73
|
4.16
|
6.1
|
|
|
1819
|
q100SW10-2
|
10
|
84.051
|
84.847
|
0.80
|
3.26
|
4.8
|
|
|
1819
|
q100SW10-3
|
10
|
87.238
|
88.035
|
0.80
|
3.32
|
4.9
|
|
|
1819
|
q100SW10-4
|
10
|
89.231
|
89.231
|
0
|
3.21
|
4.7
|
|
|
1819
|
q100SW12-5
|
12
|
30.03
|
30.03
|
0
|
3.03
|
4.5
|
|
|
1819
|
q100SW4-6
|
4
|
66.739
|
66.739
|
0
|
3.01
|
4.4
|
|
|
1819
|
q100SW4-7
|
4
|
67.808
|
67.808
|
0
|
3
|
4.4
|
|
LL
|
2018/2019
|
qLL10-2.1
|
10
|
42.768
|
51.909
|
9.14
|
8.71-10.48
|
12.3-14.6
|
| |
1819
|
qLL10-1
|
10
|
29.322
|
62.784
|
33.46
|
14.21
|
19.3
|
|
LD
|
2018/2019
|
qLD10-2.1
|
10
|
42.768
|
51.909
|
9.14
|
4.3-6.64
|
6.3-9.5
|
| |
2018/2019
|
qLD12-4.1
|
12
|
27.316
|
32.654
|
5.34
|
3.44-5.91
|
5.1-8.5
|
| |
1819
|
qLD1-1
|
1
|
3.65
|
12.708
|
9.06
|
6.98
|
10
|
| |
1819
|
qLD10-2
|
10
|
34.771
|
37.327
|
2.56
|
6.8
|
9.8
|
| |
1819
|
qLD10-3
|
10
|
42.768
|
50.744
|
7.98
|
8.1
|
11.5
|
| |
1819
|
qLD12-4
|
12
|
13.421
|
13.421
|
0
|
6.04
|
8.7
|
|
|
1819
|
qLD12-5
|
12
|
23.146
|
23.146
|
0
|
6.11
|
8.8
|
|
|
1819
|
qLD12-6
|
12
|
24.122
|
27.342
|
3.22
|
7.17
|
10.3
|
|
LA
|
2018/2019
|
qLA10-2.1
|
10
|
39.835
|
58.118
|
18.28
|
6.46-7.74
|
9.3-11
|
|
|
1819
|
qLA10-1
|
10
|
42.768
|
43.567
|
0.8
|
3.33
|
4.9
|
|
|
1819
|
qLA10-2
|
10
|
44.095
|
50.744
|
6.65
|
3.71
|
5.4
|
|
FS(SSC)
|
2019
|
qFS3-1
|
3
|
100.85
|
102.109
|
1.26
|
5.48
|
9.1
|
|
|
2019
|
qFS5-2
|
5
|
10.18
|
10.18
|
0
|
5.4
|
9.1
|
|
FF
|
2019
|
qFF10-1
|
10
|
74.412
|
76.613
|
1.2
|
4.81
|
8.1
|
*FW, fruit weight; FL, fruit length; FD, fruit diameter; FSI, fruit shape index; Nof/ec, number of fruits/end cluster; Nof/n, number of fruits/node; Nos/f, number of seeds/fruit; 100SW, 100 seed weight; LL, leaf length; LD, leaf diameter; LA, leaf area; FS (SSC), fruit sweetness (soluble solid contents); FF, fruit firmness.
{kind=link}
{kind=link}
{kind=link}
{kind=link}